Maize
Genetics Cooperation Newsletter 80. 2006.
Columbia, missouri
University of Missouri
A trans-acting factor required for non-disjunction of the B chromosome is
located distal to the TB-4Lb breakpoint on the B chromosome
--Lamb, JC; Han, F; Auger, DL; Birchler, JA
At
the second mitotic division during pollen development, the B chromosome is not
distributed to both of the daughter cells. Instead, at a high frequency one sperm cell receives both of
the sister B chromatids and the other sperm gets none. The mechanism that causes this
phenomenon, called non-disjunction, is not known.
In
a reciprocal translocation stock involving A and B chromosomes, the chromosome
with the B centromere (the B-A chromosome) is subject to non-disjunction, but
not the reciprocal translocation element containing an A centromere and the
distal portion of the B chromosome (the A-B chromosome) (Roman, Genetics
32:391-409, 1947). In order for the B-A chromosome to non-disjoin,
an A-B chromosome must be present in the same cell, suggesting that a
trans-acting factor resides near the distal tip of the B chromosome long arm (Roman, Genetics 35:132,
1950).
Translocation
TB-4Lb resulted from an interchange involving the long arm of chromosome 4 and
the B chromosome (Beckett, MNL 56:47,
1982). The breakpoint on the B chromosome is
very near the tip of the long arm so that the B-A chromosome contains almost
the entire B chromosome (Figure 1).
To determine whether
the physical location of the trans-acting factor on the B
chromosome long arm was proximal or distal to the breakpoint, we tested the
ability of the B-4Lb chromosome to undergo non-disjunction without the
reciprocal 4Lb-B chromosome.
B-4Lb
is particularly interesting because its breakpoint is distal to a small region
near the tip of the long arm of B that is enriched for the B-specific sequence (Lamb et al., Chromosoma
113:337-349, 2005)
(Figure 1), which is primarily present in and around the B centromere (Alfenito and Birchler,
Genetics 135:589-597, 1993; Jin et al., Plant Cell, 2005). This B-specific region could
be involved with the trans-acting effect on the action of the B centromere,
given their similarity in sequence.
If non-disjunction results from an interaction between the two sites of
B specific repeats, then the B-4Lb chromosome should exhibit non-disjunction in
the absence of the 4-B chromosome.
For
many B-A translocations, including TB-4Lb, pollen containing the B-A chromosome
and a normal chromosome 4 will not succeed in pollinating a tester line because
the A chromosome segment on the B-A chromosome is present in two copies,
reducing the competitive ability of the pollen (Auger and Birchler, J.
Hered. 93:42-47, 2002).
Because plants cannot be recovered that contain the B-4Lb chromosome from a
test cross using a plant with two intact copies of chromosome 4 and a B-4Lb
chromosome, we performed fluorescent in situ hybridization on pollen grains to
assay directly the ability of TB-4Lb to undergo non-disjunction.
Plants
that contained two intact copies of chromosome 4 and the B-4Lb chromosome
(called tertiary trisomics), as well as plants containing one intact chromosome
4, one B-4Lb and one 4-B chromosome (euploid heterozygotes), were selected from
progeny of a euploid heterozygote crossed as a female by a tester line. The
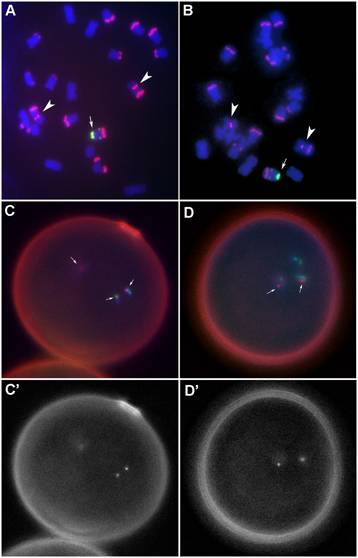
karyotype of each plant was determined by examining mitotic chromosome spreads
using a combination of FISH probes made from DNA elements specific to
chromosome 4 (Cent4), centromeres (CentC), and the B chromosome (ZmBs) (Figure
1). See Kato et al. (Proc. Natl. Acad. Sci.
USA 101:13554-13559, 2004)
for a further description of the probes.
Mature
pollen from both genotypes was fixed in 3:1 acetic acid:ethanol for 24 hours
and then stored in 70% ethanol at -20�C. Pollen was rinsed in 2XSSC and then
suspended in a probe mixture containing the B-specific element (ZmBs) and the
180 bp knob repeat in 2XSSC, 50% formamide and heated to 95C for 5 minutes.
After heating, the pollen was incubated in the probe mixture at 37C for 24
hours. The pollen was suspended in VectaShield containing DAPI to stain the
DNA, dropped onto slides, and covered with a cover slip. After waiting one hour
for the DAPI to penetrate the pollen walls, the slides were examined with a
fluorescent light microscope and images captured using a Magnafire CCD camera.
Over
100 well-labeled pollen grains from both the tertiary trisomic and the euploid
heterozygote were examined for the presence of the B-specific probe signal in
one or both sperm nuclei. The 180 bp knob probe serves as a positive control to
confirm that the probe had penetrated into and hybridized to the three pollen
nuclei. In the tertiary trisomic, about half of the pollen contained signals
from the B-specific element and in every case both sperm nuclei contained the
B-specific signal. About half of the pollen from the euploid heterozygote
contained the B-specific probe and of these, half of them showed signal in both
sperm. The remaining quarter of the total pollen had signal in only one of the
two sperm, which is an indication of non-disjunction (Figure 1).
The
pollen FISH procedure allows direct visualization of the results of B
non-disjunction in the pollen grain. This allowed us to determine that the
B-4Lb chromosome alone is incapable of non-disjunction. This result indicates
that the trans-acting factor responsible for B non-disjunction is distal to the
breakpoint of TB-4Lb and does not involve the distal ZmBs region of the B
chromosome (Figure 1A). Thus, the mechanism for B non-disjunction does not involve
interaction between the centromeric and distal B-specific sequences, and the
trans-acting factor resides at a very distal position on the chromosome.
_________________________________________________
Please
note:
Notes submitted to the Maize Genetics Cooperation Newsletter may be cited only
with consent of the authors.
Return to MNL 80 on-line index.
Return to MNL index.
Return to MaizeGDB home page
____________________________________________