NOVOSIBIRSK, RUSSIA
Russian Academy of Sciences
SURGUT, RUSSIA
Surgut State University
The fate of backcross offspring of apomeiotic maize × gamagrass hybrids
— Sokolov, VA; Kravchenko, AY; Belova, IV; Tarakanova, TK
Previously we have reported an independent genetic control of apomeiosis and parthenogenesis in diplosporous apomictic tetraploid (2n=4x=72) gamagrass (Tripsacum dactyloides — Td) (Sokolov, VA et al., MNL 74:55–57, 2000). At the same time, similar results were obtained for another diplosporous apomictic plant Erigeron annuus (Noyes, RD, Int J Plant Sci 161:1–12, 2000).
Obtaining of a single family without parthenogenesis (15 individuals, all of BIII type) among dozens of studied lines practically excluded any doubts about independent inheritance of two apomixis elements in our experiment. However, later results in aposporous apomictic plants (Ozias-Akins, P et al., Funct Integr Genomics 3:94–104, 2003) have shown non-Mendelian effects in inheritance of asexual seed reproduction and reports on genome dynamics in newly produced allopolyploids (Liu, B and Wendel, JF, Curr Genomics 3:1–17, 2002; Comai, L, Plant Mol Biol 43:387–399, 2000), which our wide hybrids also belong to, led us to the necessity of additional verification for the truth of our conclusion on the independent inheritance of apomeiosis and parthenogenesis in gamagrass and its apomictic hybrids with maize. The absence of apomixis expression in any offspring of these generations would be a sufficient proof for the independent genetic control of two basic apomictic components — apomeiosis and parthenogenesis. Besides, such a check would allow to find out the thing at which ratio of sexual : apomictic parent genomes the transfer to sexual reproduction will begin in hybrids with absence of one of asexual seed reproduction elements. According to earlier observations of apomictic maize × gamagrass hybrids, completely sexual progenies were found only after accumulation of 7 maize genomes as a minimum — 2n=9x=106 (70Zm+36Td) through BIII hybridization. In the present research we studied BC3 and BC4 of the family 363 without parthenogenesis by means of metaphase analysis of chromosome number and on a manifestation of pollinators marker character used in backcrossing. We also estimated the imprinting effect depending on the ploidy level of maize used as pollen parent. Metaphase preparations were made with standard methods, and not less than 5–7 metaphases were analysed per each plant. The number of maize genomes in hybrid was determined on the base of number of satellite maize chromosomes #6. Estimate of imprinting was carried out comparing the weight of seeds produced from hybridization with diploid and tetraploid pollinators.
Actuality of reliable confirmation for polygenic control of diplospory is connected with the necessity of working out the strategy for further research in production of asexual agricultural plants with non-segregating agronomic trait complexes. According to many experimental data, aposporous asexual seed reproduction is inherited as one big locus (Ozias-Akins, P et al., Funct Integr Genomics 3:94–104, 2003; Jessup, RW et al., Crop Sci 42:1688–1694, 2002; Stein, J et al., Theor Appl Cenet 109:186–191, 2004) whose size has not been determined yet. These results are used to ground up the possibility of genetic engineering approach to produce apomicts of any cultivated plants by transfer of the corresponding chromosome site. Based on the obtained results we support the point of view of other colleagues (Blakey, CA et al., Genome 44:222–230, 2001; Carman, JG, Biol J Linn Soc 61:51–94, 1997) about polygenic control of diplosporous apomixis and the difficulties of its transfer by present molecular-genetic tools. Thus, we believe that wide hybridization with wild related species as we performed it in maize is the most successful way of creation of apomicitic crops at the modern level of methodical base.
Seeds produced in pollination of three BC1 plants, family 363 (2n=56; 20Zm+36Td), by di- and tetraploid maize were used in our investigation. It is necessary to note that very low seed set was observed in producing BC2 hybrids, one of the reasons of which could be the imprinting effect. That is why di- and tetraploid maize was used in pollination. Theoretically, BC2 plants could be apomictic and/or initiate non reduced (2n=66 or 2n=76) or reduced (2n=38 or 2n=48) hybrids depending on the used pollinator. Alongside with this, other variants connected with abnormal chromosome distribution are not excluded. The produced 12 kernels were quite shriveled, and we did not carry out their cytogenetic analysis to avoid the loss of plants as a result of root damage in fixation of mitotic zone for metaphase analysis. Five plants grown from these seeds were similar to those of 38- (2n=20Zm+18Td) and 48- (2n=30Zm+18Td) chromosome hybrids in their habitus and developed up to flowering stage and were pollinated by di- and tetraploid maize. Very low seed set was also observed in this hybridization, and only 37 BC3 kernels were obtained from the pollination of more than 10000 florets. Out of 18 kernels planted in Novosibirsk, 14 developed to reproduction stage, and only 8 out of them produced their seeds; two hybrids had fertile pollen and seeds from selfing, and also from pollination by di- and tetraploid maize. The rest 6 plants had abnormal habitus and it was impossible to obtain progenies from them despite intensive pollination. We used other 19 kernels of this backcross in the joint project with Japan (National Institute of Livestock and Grassland Science, Nishinasuno). Eight plants developed to reproduction stage, 6 out of them had no seeds. One of two fertile plants had 36% of viable pollen. Analysis of karyotypes performed in 8 plants grown in Japan revealed variation in chromosome number from 28 to 56 (Fig. 1). Based on this one can hypothesize that the initial BC2 plants were 38- and 48-chromosome BII hybrids, and all BC3 progenies are sexual and produced from fertilization of reduced egg cells by haploid and diploid sperm. Herewith, plants with chromosome number about 30 originated from the pollination of reduced egg cells by diploid maize and, due to the imprinting effect, viable shooting kernels were only those having maize female : male genome ratio close to 2:1, respectively. Some part of backcross plants carries certain gamagrass chromosomes left from the haploid set randomly distributed at meiosis. We conclude it based on the absence of these hybrids progenies in BC3 when pollinated by tetraploid maize, again due to imprinting and a considerable deviation of parent genomes ratio in the endosperm from the required of female : male ratio equal to 2:1, respectively.
Similar observations were made also in BC3 hybrids grown in Novosibirsk. All 8 fertile plants produced the biggest seed number when pollinated by tetraploid maize and they were significantly different in seed weight compared to pollination by diploid maize. All the progenies from pollination by diploid maize with the anthocyan dominant marker were stained, i.e. they had hybrid origin.
From data mentioned above, it follows that none of the plants in BC1 progenies (2n=56, 20Zm+36Td) expressed parthenogenesis and they were not apomictic. All the BC2 plants and next generations were reduced sexual hybrids some of which had additional gamagrass chromosomes and, thus, they produced seeds due to imprinting suppression in hybridization of parent plants with different ploidy level (Sokolov, VA and Khatypova, IV, Genetika (Yugoslavia) 32:331–353, 2000). In the third backcross (BC3) all plants were also sexual and had fertile pollen that indicates complete absence of apomictic elements. We can make a conclusion that all analyzed BC1 progenies did not manifest parthenogenesis. Hence, apomeiosis and parthenogenesis in gamagrass and its apomictic hybrids with maize have independent genetic control and can segregate.
The research was carried out with the support of Russian Foundation of Basic Research, grant 03-04-49658.
Table 1. Comparative seed set and kernel weight of BC3 pollinated by diploid and tetraploid maize and by self.
| Cobs # | Number of seed rows per cob |
Number of kernels per cob |
Average kernel weight | Pollinator ploidy |
| 1 | 12 | 63 | 0.122 ± 0.010 | 2n = 4x |
| 2 | 12 | 96 | 0.144 ± 0.006 | 2n = 4x |
| 3 | 12 | 54 | 0.176 ± 0.006 | 2n = 4x |
| 4 | 10 | 41 | 0.131 ± 0.010 | self |
| 5 | 12 | 1 | 0.066 | 2n = 2x |
| 6 | 10 | 5 | 0.031 ± 0.007 | 2n = 2x |
| 7 | 12 | 34 | 0.019 ± 0.003 | 2n = 2x |
| 8 | 12 | 18 | 0.022 ± 0.006 | 2n = 2x |
| 9 | 12 | 4 | 0.034 ± 0.008 | 2n = 2x |
The differences in kernel weight at pollination by diploid and tetraploid maize are significant at P<0,001.
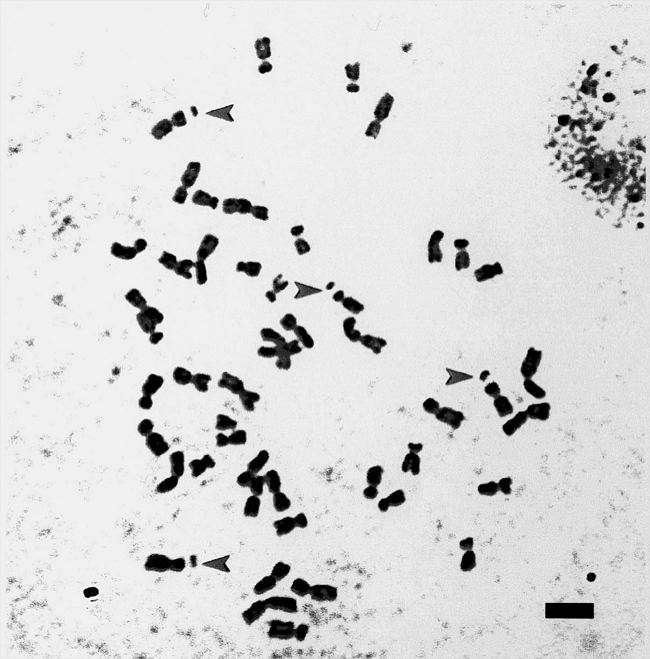
Figure 1. BC3 mitotic metaphase (2n=49). Scale 5 mm. Four maize chromosomes number 6 pointed by arrows.
Return to the MNL Volume 79 Index
Return to the index of Maize Newsletters
Return to the Maize Genome Database Page
{kind=link}