DNIEPROPETROVSK, UKRAINE
Institute of Grain Farming, Ukrainian Academy of Agrarian Sciences
Electrophoretic analysis of progeny of maize matroclinal haploids
--Dzubetskiy, BV, Satarova, TN, Cherchel, VYu, Klyavzo, SP
Intensifying the breeding process in maize requires methods for rapid establishment of homozygotic lines. Inbreeding is the most widely used method to obtain homozygotic material, but requires several years. In contrast, obtaining androgenic and matroclinal haploids and doubling their chromosomes requires only 1–2 years, and should lead to complete homozygosity.
Distinguishing matroclinal haploids through genetic markers created by S. Chase (Bot. Rev. 35:117–167, 1969) allows selection of naturally occurring haploids that appear as a result of the development of unfertilized egg cells. Crossing an original population with a line containing dominant genes of plumule coloration allows visual separation of seeds with haploid embryos from hybrid seeds.
The method of genetic markers developed in detail by E. Zabirova and O. Shatskaya (Proc. Int. Symp.: Genetics, Selection and Technology of Corn Cultivation, Krasnodar, Russia: 1999, 219–227, 1999) was used to obtain haploids. Intercrossing of different lines derived from Lancaster germplasm previously created the population Synt 1950/1952, which then became the source for obtaining haploids. The line, Krasnodarskiy Embryo Marker - 1 (ZMK-1), kindly donated by Dr. E. Zabirova and Dr. O. Shatskaya, was used as a pollinator in order to recognize haploids. In our experiment, 1266 kernels were examined after crossing with the marker line. The frequency of haploids was 1.8% of the total number of kernels observed. Only 54.8% were confirmed as haploids among the separated seeds with possible haploid embryos that had germinated in soil. Diploidization of haploids was carried out at the 3–5-leaf stage, with a 0.125% solution of colchicine, according to the technique of E. Zabirova et al. (Maize and Sorghum 4:17–19, 1996). The efficiency of diploidization was 10.5%, but only two dihaploids were self-pollinated owing to the difference in dates between tasseling and ear flowering. Thus, we obtained dihaploid (autodiploid) lines DKh 50/02 and DKh 55/02 from the initial population Synt1950/1952.
Zein, the main storage protein in corn grain, demonstrates extremely high heterogeneity, and includes up to 22 bands in its electrophoretic spectrum. Examination of the spectra of zein reveals the level of heterozygosity of many loci dispersed among several chromosomes. Of lines that are homozygous in zein-coding loci (evaluated by electrophoresis), the great majority of self-pollinated lines show no variability in morphological and agronomic characteristics. In heterozygotes, bands of both maternal and paternal forms are present in the isozyme spectrum (Asika, Poperelya, Control of genetic purity of maize hybrids by electrophoretic analysis, Methodological recommendations, Min. of Agricult., Moscow, 1991). We investigated the reduction of electrophoretic band diversity of zein components in matroclinal dihaploid lines DKh50/02 and DKh55/02 compared to the source population.
The electrophoretic analysis of zein was carried out using polyacrylamide gel with an acetic acid-glycine buffer. The acid buffer and high concentrations of urea and 2-merca-proethanol separated the storage proteins into polypeptides, each representing the product of a single gene or a cluster locus (Poperelya, Asika, Lazarev, 1991, A.C. (Patent) USSR N 16822899). For the line DKh50/02, 80 kernels resulting from the second self-pollination and 10 kernels from the first self-pollination of the line DKh55/02 were analyzed. Forty kernels of the initial population Synt 1950/1952 were analyzed.
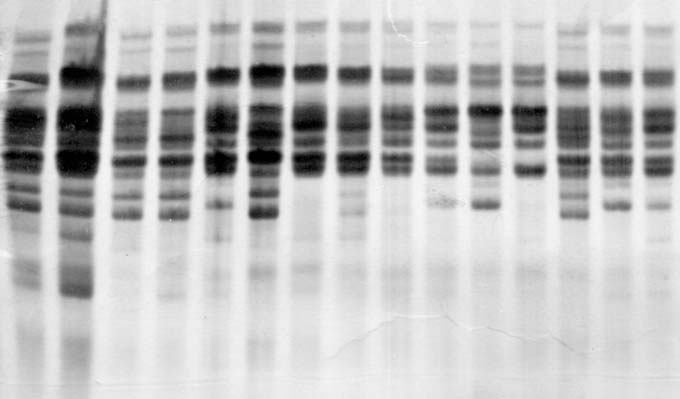
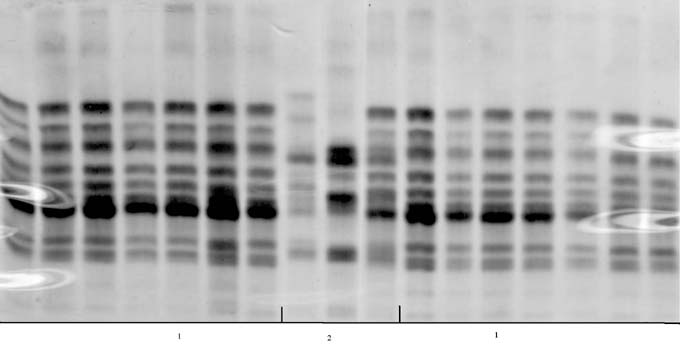
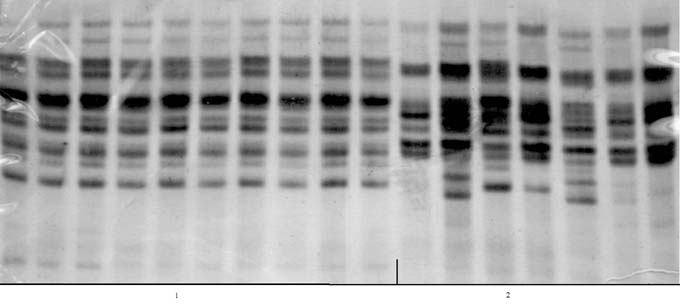
In 1950/52, there were four isozyme patterns (biotypes) containing more than one kernel (with 2,3,5 or 6 kernels each). The other 24 kernels each showed a unique pattern. This demonstrates that the initial population was highly heterogeneous for the polypeptides of zein (Fig. 1). In contrast, all the kernels of the dihaploid line DKh50/02 revealed the same isozyme pattern (Fig. 2). All the kernels of the other dihaplod line (DKh55/02) revealed a uniform isozyme pattern that was different from that of DKh50/02 (Fig. 3). The spread of some individual bands was also reduced in the dihaploid lines compared to the initial population, and some bands present in the initial population were not present in the dihaploids.
The uniformity of the protein components of zein, as the main storage protein and one with complex composition, confirms that the matroclinal haploid lines identified by the genetic marker were homozygous.
{kind=link}
{kind=link}
{kind=link}