CORVALLIS, OREGON
Oregon State University
Re-targeting of a duplicated gene product and the evolution of novel gene function
--Kulhanek, D, Keppinger, J, Rivin, C
The tetraploid ancestry of maize and related species has resulted in a genome consisting of many collinear chromosome segments carrying duplicated genes. The retention of such a high proportion of these duplicated genes is likely to derive from a variety of selection mechanisms. Our studies of the duplicate maize genes cpx1 and cpx2 suggest that they have diverged in function by a change in protein targeting that is unique to the Maydeae.
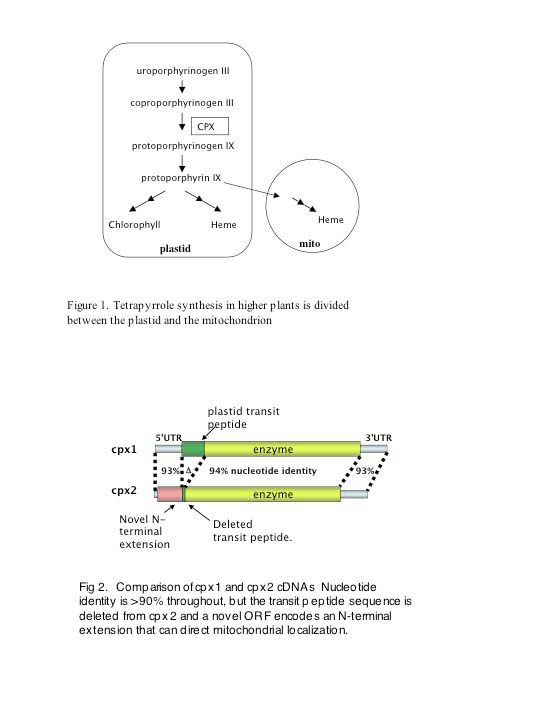
Maize cpx1 and cpx2 occupy syntenous positions on chromosomes 2 and 10 and encode almost identical enzymes for coproporphyinogen III oxidase (AKA coprogen, CPX or CPO), a critical step in the synthesis of chlorophyll and heme (Figure 1). Both genes are expressed at similar levels in all tissues. To date, CPX activity has been found exclusively in the plastids in plants. Like all other plants, the cpx1 gene encodes an enzyme with an N-terminal extension that functions as a plastid transit peptide. However, the cpx2 gene is deleted for all but a small remnant of this targeting information. At the same time, small insertions and deletions create a new contiguous 5′ ORF for cpx2 that has the features of a mitochondrial targeting peptide (Fig. 2). To test for targeting, we fused the first exon of cpx1 and cpx2 to the GFP gene and introduced the fusions biolistically into maize leaves. The cpx1-GFP localized to the chloroplasts and the cpx2-GFP localized to mitochondria (see Williamson et al., MNL 2002).
The sequenced plant genomes, rice and Arabidopsis, have only a cpx1-type gene containing a plastid targeting peptide. This is also the only type of cpx found in barley and wheat. Using PCR, we looked for the presence of the cpx2-type gene in a variety of monocots, including various Zea species, Z. mays subspecies and Tripsacum dactyloides. Both types of genes are found within the Maydeae, but we have no evidence for cpx2 outside this group of plants. The characteristic first exon of each gene was sequenced from Tripsacum and several Zea species. There was strong conservation of the cpx1 and cpx2 types of targeting information, as well as the beginning of the enzymatic region of the proteins.
The maintenance and expression of a cpx2 gene in maize and its relatives implies that it has selective value, but the role of this protein is mysterious. The substrate for CPX is thought to be solely in the plastid. A mitochondrially localized CPX2 suggests that earlier steps of heme biosynthesis may occur uniquely in the mitochondria of maize and its relatives. A null mutant for the cpx1 gene was obtained by TUSC screening with the help of Pioneer HiBred Int’l. This mutant lacks all chlorophyll and is necrotic in the light due to the accumulation of phototoxic tetrapyrrole intermediates. However, seedlings and callus cultures can be maintained indefinitely in the dark on culture media, implying a capacity to synthesize heme (and not chlorophyll) in the absence of CPX1. We have created double mutants of cpx1 and cpx2 to test whether this growth capacity in the dark depends on the cpx2 gene contributing to the production of heme. We are also looking at the effect of a cpx2 mutation on the necrotic phenotype of the cpx1 mutant in the light, to learn if CPX2 may act to scavenge and detoxify tetrapyrrole intermediates in the cytoplasm or mitochondria.
{kind=link}