Tallahassee, Florida
Florida A & M University
Florida State University
St. Paul, Minnesota
University of Minnesota
FISH analysis of retroelement distribution patterns along mitotic chromosomes.
--Bassie, YR, Onokpise, OU, Odland, WE, Bass, HW
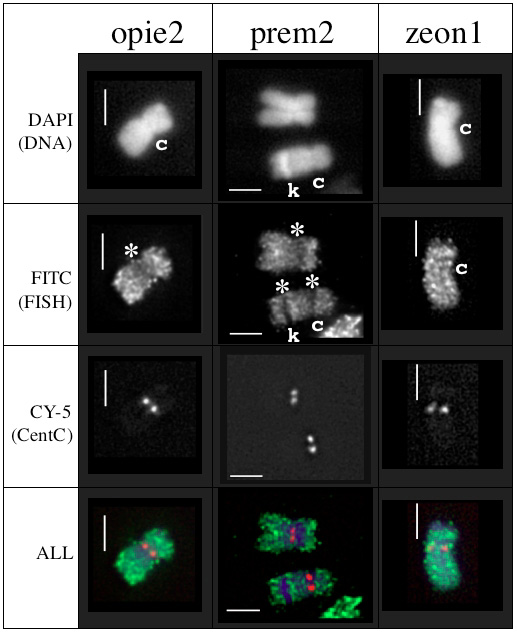
The distribution of retroelements was investigated by FISH analysis of mitotic chromosomes. PCR products specific for prem2, opie2a, and zeon1 were cloned into plasmids and used to make FISH probes following the direct-labeled procedure for chromosome painting described by Koumbaris & Bass (Plant J. 35:647–659, 2003). The chromosomes were obtained as flow-sorted mitotic root-tip chromosomes (courtesy of Dr. Arumugunathan). The slides hybridized with prem2 and zeon1 contained maize chromosome 9 from an oat-x-maize addition line. The slide hybridized with opie2a contained total maize chromosomes from B73.
Each hybridization contained two fluorescent probes — an A488-labeled probe for the retroelement (either prem2, opie2a, or zeon1) and a centromere-specific oligonucleotide CentC probe, MCCY (Koumbaris & Bass, MNL 76, 2002). Following overnight hybridization, the slides were washed, DAPI-stained, and imaged by deconvolution microscopy. Representative FISH patterns are shown in Figure 1 (wavelengths = rows; retroelement experiments = columns). As described by Mroczeck & Dawe (Genetics 165:809, 2003), retroelements often appear under-represented at centromeres (c) and knobs (k) compared to DAPI-normalized bulk chromatin on meiotic chromosomes. This pattern (Fig. 1; * indicates area of staining gap) is also clear on mitotic chromosomes hybridized with prem2 or opie2a.
In contrast, the zeon1 FISH staining pattern did not show a conspicuous reduction in signal intensity across the centromere of maize chromosome 9. The basis for these varied patterns remains unknown, but the zeon1 family may prove informative on the relationship between heterochromatin and retroelement sequences in the maize genome. In addition, understanding the distribution of retroelement families should contribute to a better understanding of the structure and evolution of the maize genome.
This work was supported in part by a fellowship to YRB from the Consortium for Plant Biotechnology Research (CBPR).
{kind=link}