Are there some species-specific differences in DNA import activity in mitochondria?
--Nepomnyaschih, DV, Koulintchenko, MV, Konstantinov, YM
It has been shown previously (MNL 64:67–68, 1990) that bacterial vector plasmids pBR322 and pBR327 can effectively penetrate into intact mitochondria of seedlings and serve as templates for DNA synthesis. Using the well-defined mitochondrial linear plasmid of 2.3 kb, we showed the existence of an active, transmembrane potential-dependent mechanism of DNA uptake into potato tuber mitochondria (Koulintchenko et al., EMBO J. 22:1245–1254, 2003). The DNA import is restricted to double-strand DNA, but has no sequence specificity. The process is most efficient with linear DNA fragments up to a few kilobase pairs. It has been concluded that DNA transport into mitochondria might represent a physiological phenomenon with some yet unknown relevance.
The aim of the present work was to investigate the potential differences in DNA import activity between mitochondria from maize seedling and potato tuber mitochondria.
Mitochondria were isolated from potato (Solanum tuberosum) tubers as described (Koulintchenko et al., EMBO J. 22:1245–1254, 2003). Maize mitochondria were isolated from 4-day-old etiolated seedlings of hybrid VIR42 MV by the standard method of differential centrifugation. The substrate DNA used for mitochondrial import assays was the orf1 fragment of 2.3 kb linear plasmid from maize (Leon et al., 1989; DDBJ/EMBL/GeneBank accession No. X13704). To obtain the radioactive linear fragment, 50 ng of unlabeled PCR product and the primers (ATGACAAAGACATATAGAATATCC / TTATTCCT TTTCGCGCTGCATT) were used for a single PCR cycle in which a 10 min elongation in an unlabeled dATP-deprived reaction medium containing 100 mCi of [α-32 P]dATP (3000 Ci/mmole) per 50 μl was followed by the addition of 0.2 mM unlabeled dATP, and a further 5 min elongation.
Standard mitochondrial import of DNA was carried out in a solution (import buffer) containing 40 mM potassium phosphate, pH 7.0, and 0.4 M sucrose. The samples (200 μl) containing 1–5 ng of 32P-labeled DNA and purified mitochondria (ca 300 μg of protein) were incubated at 25 °C for 45 or 90 min under mild shaking. Following addition of 200 μg of DNase I and 10 mM MgCl2, the incubation was continued for 20 min in the same conditions. Mitochondria were subsequently washed three times by resuspension in 1 ml of solution containing 10 mM potassium phosphate, 300 mM sucrose, 10 mM EDTA, 10 mM EGTA, 0.1% (w/v) BSA, 5 mM glycine, pH 7.5, and centrifuged for 5 min at 10 000 g. The final pellets were extracted with one volume of buffer containing 10 mM Tris-HCl, 1 mM EDTA, 1% (w/v) SDS, pH 7.5, and one volume of phenol. The nucleic acids recovered in the aqueous phase were ethanol-precipitated, fractionated by electrophoresis on a 1% (w/v) agarose gel and transferred onto a nylon membrane (Hybond N+, Amersham Biosciences) for autoradiography.
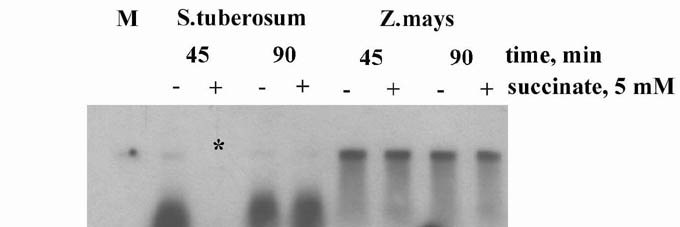
Figure 1 shows the uptake of 2.3 kb maize mitochondrial plasmid DNA by potato tuber and maize seedling intact mitochondria. Under the conditions used, maize mitochondria import DNA much more actively in comparison with potato mitochondria. To test the functional state of both potato and maize mitochondria, we also measured their respiratory control ratio (RCR), i.e. the ratio between the respiration rate of the mitochondria upon addition of ADP (150 μ) (state 3 respiration) and the respiration rate in the absence of additional ADP (state 4 respiration). RCR was rather high (3–4) with succinate-supported respiration for both plant species mitochondria. There was no detectable influence of succinate on DNA import activity (Fig. 1). The possibility that mitochondria use some endogeneous respiration substrates (like long chain fatty acids or others) under these conditions couldn’t be excluded. The differences in DNA import activity of potato and maize mitochondria might be explained by the theory that the functional state of plant mitochondria may somehow affect DNA incorporation into organelles. Further experiments are required to clarify the question of whether some species-specific differences exist in mitochondrial DNA import activity depending on plant developmental stage, tissue specialization (storage or meristematic), etc. As a whole, the studies on DNA import activity in mitochondria in a number of plant species could result in new mitochondrial transformation strategies to improve some agronomically important traits encoded by the mitochondrial genome.
Figure 1. Uptake of DNA in potato (Solanum tuberosum) and corn (Zea mays) freshly isolated intact mitochondria.
*Line where DNA wasn’t loaded.
{kind=link}