HAMBURG, GERMANY
University of Hamburg
The large second intron of the MADS box transcription factor gene ZmMADS2 is transcribed in maize tissues where the protein seems not to be required
--Dresselhaus, T, Schreiber, DN
We are interested in the function of MADS box transcription factor genes in both male and female gametophytes (pollen and embryo sac, respectively; see also Schreiber, Dresselhaus, Flowering Newsl. 35:8–16, 2003) and have cloned 7149 bp of the maize MADS box gene ZmMADS2 (GenBank accession # AY227363). ZmMADS2 consists of eight exons (E1-E8; see Figure 1) and encodes a class II MADS domain protein of the MIKC-type. This type of MADS domain protein contains an N-terminal MADS box (M: in ZmMADS2 encoded by exon 1), followed by an intervening region (I: in ZmMADS2 encoded by exon 2), a keratine-like box (K: in ZmMADS2 encoded by exons 3–5) and a highly variable C-terminus (C: in ZmMADS2 encoded by exons 6–8). The length of the transcript is around 1.3 kb and the size of the encoded protein is 240 aa.
ZmMADS2 was previously reported to be specifically expressed in maize pollen and during maize pollen tube growth (Heuer et al., Sex. Plant Reprod. 13:21–27, 2000). A more detailed expression study revealed that ZmMADS2 is also weakly expressed in root tips and strongly expressed in endothecium and connective tissues of the anther at one day before dehiscence, and in mature pollen after dehiscence. Development of both anthers and pollen was arrested at one day before dehiscence in transgenic plants expressing the 3′ part of the ZmMADS2-cDNA in antisense orientation, indicating that the protein is required for anther dehiscence and pollen maturation. High amounts of a ZmMADS2-GFP fusion protein accumulate in degenerating nuclei of endothecial and connective cells of the anther, but could not be detected in germinated pollen tubes (Schreiber et al., Plant Physiol. 134:1069–1079, 2004).
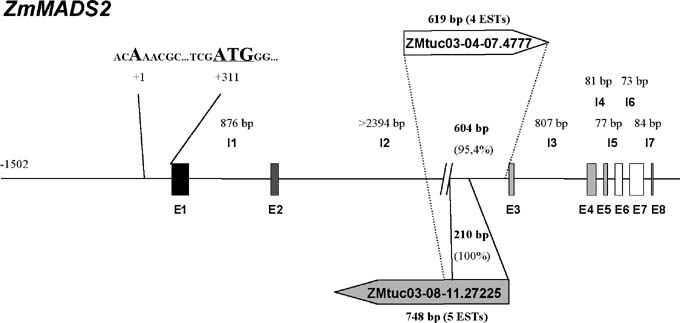
Here, we report about the second intron of the ZmMADS2 gene. Concerning MADS box genes from other plant species, this intron, with more than 2.4 kbp, is unusually large. After obtaining 2394 bp, we have stopped sequencing the second intron. The isolation of independent clones from a genomic library of the maize inbred line A188 suggests that the intron is even much bigger. None of the genomic clones isolated after screening a genomic library with the 3′ UTR of ZmMADS2 contained the 5′ UTR, although the clones were relatively large in size with a length varying between 11 and 23 kbp. Instead, we found that the 3′ part of the second intron is transcribed in various maize tissues. One end of TUC (tentative unique contig) ZMtuc03-08-11.27225 (748 bp), a contig of five ESTs (expressed sequence tags), is, over a length of 210 bp, 100% identical to the 3′ end of the second intron of ZmMADS2 (Figure 1). Adding the remaining 538 bp of ZMtuc03-08-11.27225 to the intron sequence of I2 increases the size of the intron to almost 3 kbp, although we assume that the intron is even much larger. Three of five ESTs were obtained from roots and thus overlap with the expression of ZmMADS2, while the two remaining ESTs were isolated from seedlings and the cell division part of the 6th leaf, thus tissues where ZmMADS2 transcript could not be detected. It is unlikely, that the promoter of ZmMADS2, which is located a few kbp upstream of the 3′ end of I2, influences transcription from this region. It is more likely that promoter(-like) elements in I2 are responsible for the expression. Interestingly, ZMtuc03-08-11.27225 does not contain an obvious open reading frame (ORF) and thus may not encode a protein, but instead be required for processing and stability of the ZmMADS2 transcript. Transgenic maize plants have shown that ZmMADS2 is required for anther and pollen maturation, but neither for root development nor for cell division, where low or no ZmMADS2 transcript amounts could be detected. From the expression of ZMtuc03-08-11.27225 in these tissues, we suggest that the intron-transcript could be involved in ZmMADS2 transcript stability/degradation with the aim of silencing ZmMADS2 activity in those tissues where the protein is not required.
ZmMADS2 is not a single copy gene. Using the 3′ UTR as a probe, we obtained at least three bands after digesting DNA of the inbred line A188 with various restriction enzymes (data not shown). This could be one explanation for the finding that we identified a second TUC, ZMtuc03-04-07.4777 (619 bp), transcribed from the intron of a MADS box gene allelic to ZmMADS2. Like ZMtuc03-08-11.27225, ZMtuc03-04-07.4777 does not contain an obvious ORF and thus might also be involved in transcript stability/gene regulation. Sequence identity between ZMtuc03-04-07.4777 and the 3′ end of I2 of ZmMADS2 as well as with one end of ZMtuc03-08-11.27225 is almost 96% over the whole contig. All ESTs of ZMtuc03-04-07.4777 were obtained from mixed BMS (Black Mexican Sweet) tissue and suspension cultures and therefore could also represent a DNA polymorphism. The genomic sequence of ZmMADS2, as well as the full length cDNA, were obtained from other lines, namely inbred line A188, and the ESTs of ZMtuc03-08-11.27225 were from cultivars W23 and F2.
Figure 1. Genomic structure of the MADS box transcription factor gene ZmMADS2. The ZmMADS2 gene consists of eight exons (E1 – E8) and seven introns (I1 – I7). The second intron (I2) is unusually large. The size of all seven introns is indicated. The MADS box (M) is encoded by exon 1 (black box), the intervening region (I) by exon 2 (dark grey box), the keratine-like box (K) by exons 3–5 (light grey boxes) and the highly variable C-terminus (C) by exons 6–8 (white boxes). The ATG START codon is located 311 bp downstream of the transcription start point. The 3′ region of the second intron (I2) is transcribed in various maize tissues (see text) as indicated by the two TUCs (tentative unique contigs), each represented by four and five ESTs, respectively.
{kind=link}