Evaluation of seedling growth of maize inbreds (Zea mays ssp. mays) selected from Andean-Patagonian populations for cold tolerance
--Huarte, HR, Jatimliansky, JR, Molina, MC
Maize is considered a cold susceptible crop with relatively high optimum temperatures for germination, development and dry matter accumulation (Miedema, P., Adv. Agron. 35:93–129, 1982). During the early vegetative stage, plants are susceptible to stress by low temperatures. Actual agronomic practices such as no-tillage and earliest sowing date during the growing season, increase the probability and extent of damage by cold.
The no-tillage system has spread to a great number of cultivation areas, and when this practice is used, it is very common to record soil temperatures lower than those observed under the moldboard plowing system, due to the presence of stubble on the soil surface (Griffith, DR, et al, Agron. Journal 65:321–326, 1973). A lower soil temperature increases the time to seedling emergence and consequently the risk of losses by pathogen attack (Shaw, RH, Corn and Corn Improvement, American Society of Agronomy, Madison Wisconsin, pp. 609–638, 1988), and weed competence is also greater (Olson, RA and Sander, DH, Corn and Corn Improvement, American Society of Agronomy, Agronomy Monograph 18, Madison Wisconsin, pp. 639–686, 1988). Although early sowing dates maximize the duration of the growing stage and promote yield increase (Laurer, JG, et al., Agron. Journal 91:834–839, 1999), they also increase the possibility that germination and early growing stages occur at sub-optimal temperatures. The situation described above is enough to show the need for developing maize germplasm tolerant to low temperatures during crop establishment (Lee, EA, et al., Crop Sci. 42:1919–1929, 2002). Brandolini et al. (Brandolini, A, et al., Euphytica 111:33–41, 2000), stated that tolerance to cold temperature stress could be found in exotic germplasm, for example, that of the Andean region. Therefore, the objective of this work was to compare the seedling growth of several germplasms at 10 °C (stress temperature) and at 25 °C (control temperature).
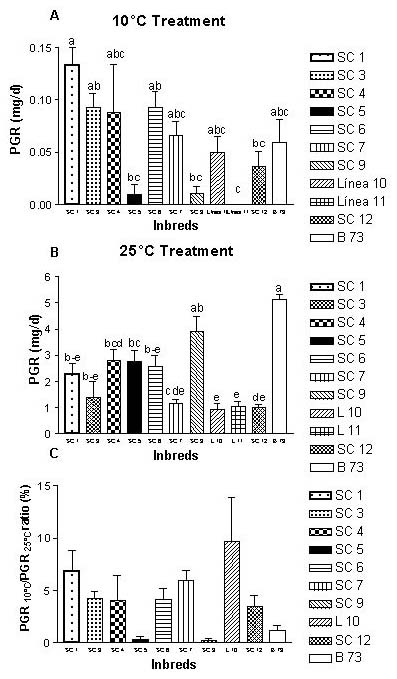
Six short cycle populations from NW Argentine Patagonia and Xth region of Chile (SC1/ SC7), a mid cycle (SC9) and a long vegetative cycle (SC12) both from temperate origins were used. As controls, three cold tolerant inbreds were used, one introduced from the USA (B73) and the remainder introduced from France (L10 and L11).
Plant growth rates at 10 °C (PGR10), at 25 °C (PGR25) and their ratio (PGR10/PGR25) were evaluated. The design was a DCA with five replications for both stress (10 °C ± 1 °C) and control treatments (25 °C ± 1 °C). Twenty-five seeds placed in nine cm Petri dishes on two discs of filter paper composed each plot. Seeds were hydrated with six ml of distilled water and re-hydrated when necessary. Incubation took place in darkness for twenty-eight days (stress treatment) and seven days (control treatment). Each germinated seed was transferred from the Petri dish to a plastic tray until the end of the experiment. At that time, the epicotyl and the radicle were excised and dried in an oven at 80 °C for 24 h, and weighed to obtain the dry matter per plot. Data were subjected to analysis of variance (ANOVA). If significant differences were detected, treatment means were separated using HSD Tukey’s test (α=0.05).
PGR10 for SC1 was greater than those of the testers (P<0.05)(Figure 1A). On the other hand, PGR25 for B73 was higher than those scored for Andean inbreds (P<0.05)(Figure 1B). PGR10/PGR25 ratios for the Andean inbreds showed a good performance. However, none of them exceeded L10 tester ratios (Figure 1C). Results suggest that some Andean-Patagonian inbreds are promising materials for developing new cold tolerant hybrids.
{kind=link}