Genomic affinities between maize and Zea perennis using classical and molecular cytogenetics (GISH-FISH)
--Gonzalez, G, Confalonieri, V, Naranjo, CA, Comas, C, Poggio, L
Zea mays ssp. mays (maize) is an allotetraploid (2n=20), and its genomic formula according to Naranjo et al. (1990) is AmAmBmBm. Zea perennis is an alloautooctoploid (2n=40) with a genomic formula ApApA′pA′p Bp1Bp1Bp2Bp2. These taxa, as well as the rest of the members of the genus Zea, are segmental cryptic polyploids, with diploidized behavior and basic number x=5 (Naranjo et al., 1990 Acad. Nac. Cs. Ex. Fís. y Nat., Bs. As. 5:43–53).
The aim of this work was to discover the homology between maize and Zea perennis. For this reason, their genomic affinities were analyzed through: a) The meiotic behavior of both parents (maize and Zea perennis) and their artificial hybrid (2n=30), through classical cytogenetics. b) The molecular affinities, at the medium and highly repetitive DNA level by genomic in situ hybridization (GISH). c) The physical mapping of the ribosomal sequence 45S (pTa71) using FISH.
The materials used were Zea perennis (from the location Piedra Ancha) and Zea mays ssp. mays (cv. 6482), both grown at the Instituto Fitotécnico Santa Catalina (Llavallol, 35 °S, 58 °W).
a) Classical cytogenetic studies: In order to know the meiotic behavior in metaphase I, chromosome preparations were made using young anthers from maize, Zea perennis and its F1 hybrids, which were dyed with acetic hematoxilin (2%).
This study showed a regular pairing of the chromosomes in maize, and the more frequent meiotic configuration was 10 II. The meiosis of Zea perennis showed a configuration of 5 IV + 10 II in most of the cells analyzed. In the F1 hybrid Zea perennis × Zea mays ssp. mays (2n=30), whose proposed genomic formula is AmApAp′Bp1Bp2Bm (Naranjo et al., 1990), the more frequent meiotic configuration was 5 III + 5 II + 5 I.
b) GISH studies:
b.1) Metaphase maize chromosomes were hybridized with labeled total genomic DNA from Zea perennis. In this experiment, a scattered signal of hybridization was observed all over the chromosomes except for two pairs, the metacentric chromosome pairs I and II (Figure 1), which show weak hybridization signal. This showed that these maize chromosomes have some sequences that are not shared with Zea perennis genome. In this experiment, it was also observed that the heterochromatic knobs of maize (DAPI+ bands), composed of a 180 bp sequence highly tandem repeated, do not show a hybridization signal with the genomic DNA of Zea perennis.
b.2) A hybridization experiment using labeled genomic DNA of maize and non-labeled genomic DNA as blocking in a proportion of 30:1 was carried out on interphasic and metaphasic maize cells. This experiment showed weak signals of hybridization on all the chromosomes except at the knobs zones (DAPI +) which exhibited an intense hybridization signal. This confirms that Zea perennis lacks the 180 pb sequence which is constitutive of maize knobs. This fact had already been observed in previous experiments of Southern-Blot and GISH (Poggio et al., 1999, Genome 42: 993–1000).
b.3) The metaphasic chromosomes of Zea perennis were hybridized with the labeled genomic DNA of maize. In this experiment, signals of dispersed hybridization throughout all the chromosomes were observed and none of them displayed a pattern of differential hybridization.
b.4) Meiotic cells (metaphase I) of the artificial F1 hybrid Zea mays ssp. mays × Zea perennis (2n=30) were hybridized with the total genomic DNA of maize. In this case, it was observed that the chromosomes that remained univalents at the meiotic configurations described above belong to maize (since they show a strong hybridization signal), and the bivalent chromosomes correspond to Zea perennis (as they show a weak hybridization signal).
These results confirm the origin of the meiotic configurations postulated in previous works.
c) FISH studies: Using the ribosomal sequence 45S of wheat (pTa71) as a probe.
c.1) This probe was hybridized on chromosomes of maize and two signals of hybridization were observed on the short arms of chromosome pair number 6.
c.2) Then, the probe was hybridized on metaphasic chromosomes of Zea perennis. In this case, four signals of hybridization were detected.
c.3) Finally, metaphases I of the artificial F1 hybrid Zea perennis x Zea mays ssp. mays (2n=30) were hybridized with the probe pTa71. This experiment showed three signals of hybridization which, in most of the cells, were located on a trivalent; whereas in the rest of the cells, two signals were located on a bivalent and only one signal on a univalent. These results indicate that the 45S ribosomic DNA sequence is present in the genomes that show the greatest degree of pairing during meiosis.
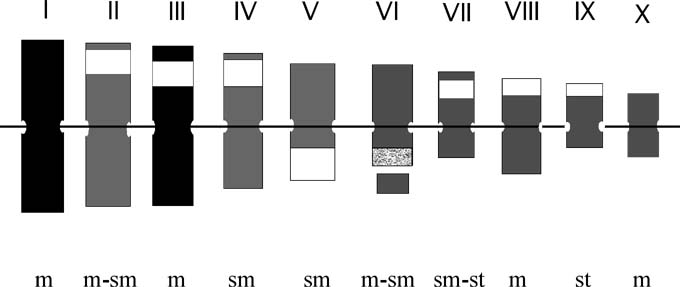
The results obtained by GISH and FISH were summarized through an idiogram of maize which shows those chromosomes with less molecular affinity with Zea perennis, the location of knobs and the physical mapping of the 45S ribosomal sequence (Figure 1).
These results allow us to conclude that an important divergence exists between the genomes of Zea mays ssp. mays and Zea perennis because of: a) the lack of hybridization observed on four chromosomes of maize (pairs I ad 2) when hybridized with Zea perennis DNA; b) the dispersed hybridization signals of the genomic maize DNA on Zea perennis chromosomes; and c) the lack of the 180 pb sequence of knobs in Zea perennis.
The classical and molecular cytogenetic studies (GISH-FISH) done on the F1 hybrid Zea perennis x Zea mays ssp. mays allow us to conclude that: a) The univalents come from maize and the bivalents from Zea perennis (by GISH); b) the fact that the 45S ribosomal DNA hybridized, in most of the cases, on a trivalent demonstrates that they are formed by two chromosomes of Zea perennis and a chromosome of maize. In addition, this result indicates that this sequence is located in the genomes with a greater degree of pairing in the hybrid (named genome A).
All the results indicate that Zea perennis (2n=40) is an autoalloctoploid in which one of its genomes would have been more compatible to the ancestor of the present maize. These results and the most frequent meiotic configurations observed in the hybrid, confirm the genomic formulas postulated in previous works (Naranjo et al., 1990; Poggio et al., 2000 Genet. Mol. Biol. 23:1021–1027) (Figure 2).
Figure 1. Idiogram of Zea mays ssp. mays (cv 6482) showing that the chromosome pairs I and II show the lowest molecular homology with Zea perennis (in black). White bands are knob locations. The dotted band is the location of the 45S ribosomal sequence (pTa71). m: metacentric. sm: submetacentric. st: subtelocentric.
Figure 2. The most frequent meiotic configurations observed in Zea mays ssp. mays, Zea perennis and their F1 artificial hybrid. Arches show the more frequent meiotic associations. Asterisks indicate the genomes that possess the 45S (pTa71) ribosomal sequence. III: trivalent; II: bivalent; I: univalent.
{kind=link}
{kind=link}