Cold adaptation induction of AOX isophorm in maize
--Borovskii, G, Lukina, E, Korotaeva, N, Stupnikova, I, Antipina, A, Truhin, A, Popov V
Higher plants have two mitochondrial electron transport pathways from ubiquinone to oxygen. One is an electron transfer through the cytochrome pathway coupled to ATP production and inhibited by cyanide. The alternative pathway branches from the cytochrome pathway and donates electrons to oxygen directly by alternative oxidase (AOX) (Moore and Siedow, Biochim. Biophys. Acta. 1059:121–140, 1991). The alternative pathway is not coupled with ATP synthesis, but can be induced in response to stress or inhibition of the main electron transfer pathway. When plants are exposed to low temperatures, they produce reactive oxygen species (ROS) such as superoxide and hydrogen peroxide (Prasad et al., Plant Cell. 6:65–74, 1994). ROS damages membranes and other cell components. It has generally been proposed that increased AOX activity could be used to help maintain normal levels of metabolites and to reduce levels of reactive oxygen species (ROS) during the cold. In maize the part of electron partitioning to the alternative pathway was significantly increased under low temperatures, though it may not correlate with tolerance to cold (Ribas-Carbo et al., Plant Physiol. 122:199–204, 2000). A nuclear gene encodes alternative oxidase. The genes of AOX consist of a small multigene family. Three members were found in soybean (Whelan et al., Planta 198:197–201, 1996), rice (Ito et al., Gene 203:121–129, 1997; Saika et al., Genes. Genet. Syst. 77:31–38, 2002) and four members in Arabidopsis (Saisho et al., Plant Mol. Biol. 35:585-596, 1997) and mango (Considine et al., Plant Physiol. 2001 126:1619–1629). In maize a set of isophorms of AOX have also been demonstrated (Karpova et al., Plant Cell. 14:3271–3284, 2002).
The alternative oxidase protein levels in the mitochondria were studied after short cold stress at 2 °C, as well as after adaptation during 7 days at 12 °C. Four day etiolated maize seedlings grown at 20 °C were used as control. Mitochondria were isolated and purified as described earlier (Borovskii et al., BMC Plant Biol. 2:5, 2002). The expression of AOX proteins was performed by immunoblotting with AOX specific antibodies (kindly provided by Dr. T. E. Elthon, University of Nebraska).
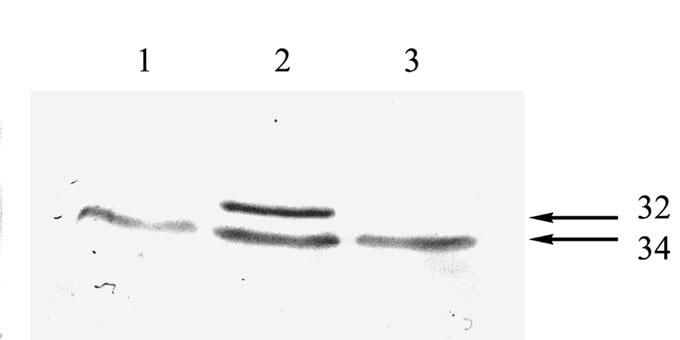
Immunoblotting revealed that the level of AOX protein is raised under low temperature adaptation (Fig. 1). It is easy to see that most of the total AOX level increased on account of augmentation of the content of the 34 kD isophorm. The level of 32 kD isophorm characteristic for control seedling mitochondria was not increased. The differential increase of the 34 kD isophorm level made us suspect that these proteins are independently regulated, and may have specific and different functions during adaptation of maize to cold temperature. A similar situation appeared in rice, where only the 34 kD isophorm of AOX had a correlation with low temperature tolerance and the 32 kD did not (Abe et al., FEBS Lett. 527:181–185, 2002). It has also been reported that the capacity of the alternative pathway is greater in winter wheat than in spring wheat (McCaig and Hill, Can. J. Bot. 55:549–555, 1977). Alternatively in maize, the alternative pathway was significantly increased in the more chilling-sensitive variety (Ribas-Carbo et al., Plant Physiol. 122:199–204, 2000). The exact role of AOX and other energy dissipative pathways, such as external and internal rotenone-insensitive dehydrogenases or uncoupling protein (UCP) activity during stress, still need to be defined.
This work was supported by the Russian Fund of Basic Research (project 02-04-48599).
Figure 1. Western analysis of maize mitochondrial AOX proteins. Mitochondria were isolated from the control (1), 7 days cold (12 °C) acclimated (2) and cold (2 hours at 2 °C) stressed (3) seedlings. Molecular weights of AOX isophorms are indicated.
{kind=link}