Isolation of a redox dependent mitochondrial DNA binding protein which binds specifically to the coxI promoter
--Tarasenko, VI, Subota, IY, Kobzev, VF, Konstantinov, YM
Although the plant mitochondrial promoter sequences are relatively well characterized, at the current time little is known about trans-factors controlling expression of the mitochondrial genome. There is only one report on isolation of mitochondrial DNA binding protein from a monocotyledon plant (wheat). This protein binds to the coxII gene upstream region and seems to be involved in transcription initiation (Ikeda and Gray, Mol. Cell. Biol. 19:8113–8122, 1999). In yeast and animal mitochondria, activity of a single transcription factor is sufficient for accurate initiation of transcription of all genes (Jang and Jaehning, J. Biol. Chem. 266:22671–22677, 1991; Fisher et al., Mol. Cell. Biol. 8:3496–3504, 1988). The authors proposed that isolated wheat protein also plays a role of such an universal transcription factor. On the other hand, the existence of multiple gene-specific mitochondrial transcription factors has been proposed on the basis of specific dependence of coxII expression in Zea perennis on the nuclear MCT locus (Newton et al., EMBO J. 14:585, 1995). Nothing is known about physiological conditions that can influence the activity of mitochondrial DNA binding proteins.
Both our research group (Konstantinov et al., Biochem. Mol. Biol. Intern. 36:319–326, 1995) and other investigators (Wilson et al., Eur J. Biochem. 242:81–85, 1996) have demonstrated the existence of redox regulation of transcription in plant mitochondria. An activation of RNA synthesis in organello in maize under oxidizing conditions and its suppression under reducing conditions was detected. Now we are trying to identify protein factors that can mediate this redox regulation. Previously, we have shown redox dependence of mitochondrial DNA topoisomerase I activity (Konstantinov and Tarasenko, MNL 73:39–40, 1999). In this letter, we report partial purification of a DNA binding protein that binds specifically to the coxI promoter region and also possesses properties of a redox dependent protein.
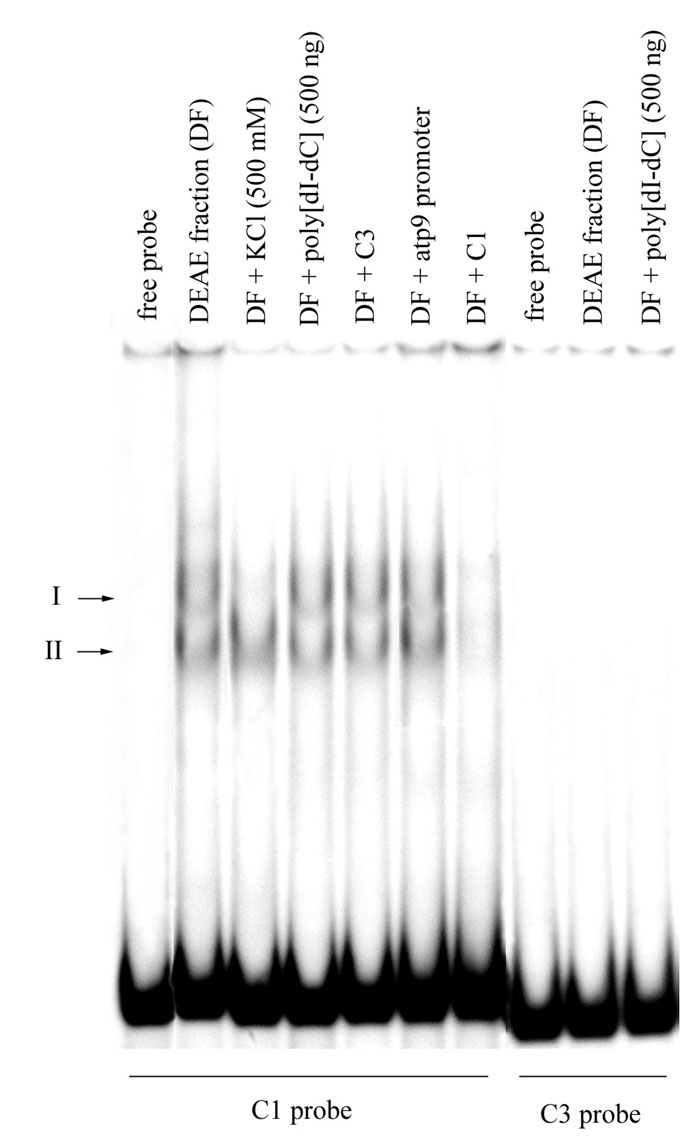
The mitochondria were isolated from 4-day-old etiolated seedlings of hybrid VIR42 MV and lysed by high salt and Triton X-100. The supernatant was applied on a DEAE-Toyopearl chromatography column. EMSA was carried out as described by Kagoshima et al. (J. Biol. Chem. 271:33074–33082, 1996) with minor modifications. We used a PCR-generated 230 bp EMSA probe containing the upstream region of the coxI gene (C1). This region includes two sites of transcription initiation (-326 and -231 relative to ATG). Another probe used (C3) contained a similar region of the coxIII gene (210 bp) which also comprises two initiation sites (-360 and -320 relative to ATG). The reason for use of such probes was the possibility that some additional motifs located upstream or downstream of the core promoter sequence may be important for specific binding.
Some fractions eluted from DEAE-Toyopearl contained activity which formed two complexes in EMSA when C1 was used as a probe (Fig. 1). These retardation complexes were stable in high salt and in the presence of non-specific competitor DNA, demonstrating the specificity of the interaction. Surprisingly, we have not detected any DNA binding activity when the C3 probe was used instead of C1. In another assay, the presence of unlabeled C3 probe, as well as a 34 bp oligonucleotide contained the atp9 promoter core sequence in 50-fold excess in retardation reaction of DEAE fractions with labeled C1, did not affect the efficiency of DNA binding. These data strongly suggest that isolated protein demonstrates specific binding to the promoter region of the coxI gene, but doesn’t possess an ability to bind to all mitochondrial promoter sequences. Based on this fact, it is possible to speculate that in maize mitochondria, unlike the situation existing in animal and yeast organelles, there is more than one transcriptional factor, and transcriptional regulation might be gene-specific.
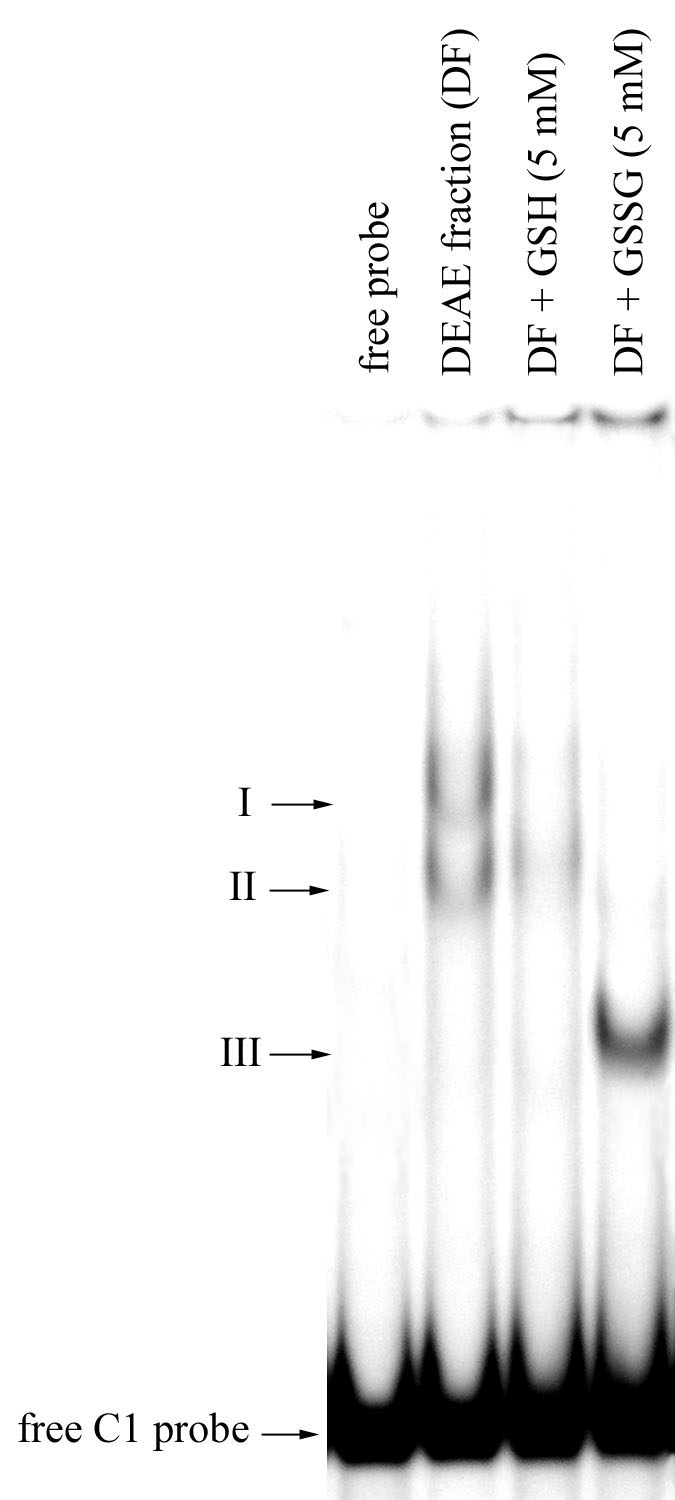
In further experiments we tested the influence of reduced (GSH) and oxidized (GSSG) glutathione on DNA binding activity of the protein (Fig. 2). Some decrease of DNA binding activity was detected in the presence of GSH. Addition of GSSG caused significant changes in the DNA binding pattern. A faster migrating band (Fig. 2, III) emerged, whereas two bands detected in the absence of the agent (Fig. 2, I and II) disappeared. Moreover, intensity of this newly formed retardation complex was higher than that of two complexes detected when GSSG was omitted. Such DNA binding modulation could be explained by changes in the protein-DNA complex conformation caused by reduction/oxidation of cysteine residues in the protein molecule.
In conclusion, we have shown that reduced and oxidized forms of glutathione modulate the activity of the mitochondrial DNA binding protein that specifically recognizes the coxI promoter region. Such modulation potentially could play an important role in regulation of expression of mitochondrially encoded proteins, especially of respiratory chain components, in response to changes in the redox state of the chain. Additional investigations are needed to verify this hypothesis. Further purification and biochemical characterization of the protein is underway in our laboratory.
Financial support from the Russian Foundation for Basic Research (Grant No. 01-04-48162) is acknowledged.
Figure 1. DNA binding activity exhibited by protein fractions isolated by chromatography on DEAE-Toyopearl (DF, eluted with 0.4 M KCl). Competition study using non-specific DNA competitor and probes containing mitochondrial promoter sequences. C1 — upstream region of the coxI gene (230 bp); C3 — upstream region of the coxIII gene (210 bp); atp9 — core promoter of the atp9 gene (34 bp, -288 relative to ATG). Unlabeled competitors were present in the reactions in a 50-fold excess relative to the labeled probe. Arrows indicate the positions of two DNA-protein complexes and the free radiolabeled C1 and C3 probes.
Figure 2. The effect of reduced and oxidized glutathione on DNA binding activity recovered in DEAE-Toyopearl protein fractions. Arrows indicate the positions of three DNA-protein complexes and the free radiolabeled C1 probe.
{kind=link}
{kind=link}