IRKUTSK, RUSSIA
Institute of Plant Physiology and Biochemistry
NOVOSIBIRSK, RUSSIA
Institute of Cytology and Genetics
The effects of the glutathione redox system on DNA binding activity of nuclear DNA topoisomerase I
--Subota, IY, Katyshev, AI, Tarasenko, VI, Senghenko, LP, Kobzev, VF, Konstantinov, YM
Redox regulation of DNA binding activity of transcription factors involving modification of conservative cysteine residues in the protein molecule is considered to be one of the key mechanisms of gene expression control (Martin et al., J. Biol. Chem. 271:25617–25623, 1996; Akamatsu et al., J. Biol. Chem. 272:14497–14500, 1997). We have previously shown that DNA topoisomerase I (topo I) from carrot mitochondria, and probably the enzymes from mitochondria of other plant species, can be regarded as redox sensitive enzymes. Existence of evolutionary conservative cysteine residues which could potentially form disulfide bridges in functionally important domains of carrot protein was demonstrated (Konstantinov et al., Proc. Russian Acad. Sci. 377:263–265, 2001). Hence, mitochondrial topo I is a very plausible candidate for involvement in the mechanism of redox control of expression in the organelles. Recent data showing influence of redox conditions on DNA binding of maize mitochondrial topo I (MNL 77:37–38, 2003) also point to the possibility of complex participation of the enzyme in gene expression regulation. It is still unclear if the demonstrated redox dependence of DNA binding activity of mitochondrial topo I could be a common characteristic of plant enzymes from different cellular locations. In principle, orchestrated redox modulation of nuclear and mitochondrial topo I activities could be one of the ways to coordinate expression of mitochondrially located proteins encoded in the nucleus and mitochondria. To verify the hypothesis about the existence of a mechanism of redox control of nuclear topo I activity we have studied the influence of such physiological redox agents as glutathione on the DNA binding activity of nuclear topo I.
The nuclei were isolated from 4-day-old etiolated maize seedlings of hybrid VIR42 MV as described by Mascarenhas et al. (Methods in Enzymology 31:558–565, 1974) with some modifications. The method of topo I isolation was the same as described earlier (MNL 74:33, 2000) with the additional step of purification on non-denaturing PAGE. EMSA was carried out as described (Ikeda and Gray, Mol. Cell. Biol. 19:8113–8122, 1999) with minor modifications. Assay employed double stranded DNA probes containing p53 and atp9 gene promoter fragments. Oxidized and reduced glutathione were used in the experiments in 5 mM concentration.
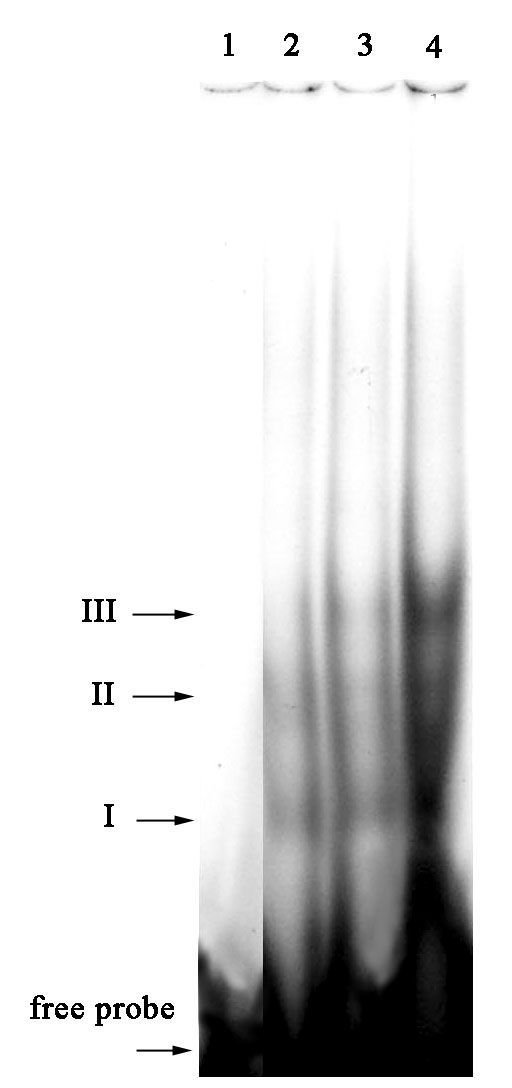
Figure 1 shows the influence of two forms of glutathione on nuclear topo I DNA binding activity towards the probe containing a fragment of the p53 gene promoter sequence. The presence of oxidized glutathione (GSSG) caused a significant increase in DNA binding (Fig. 1, I-III). However, addition of the reduced form of glutathione (GSH) did not result in any detectable effect.
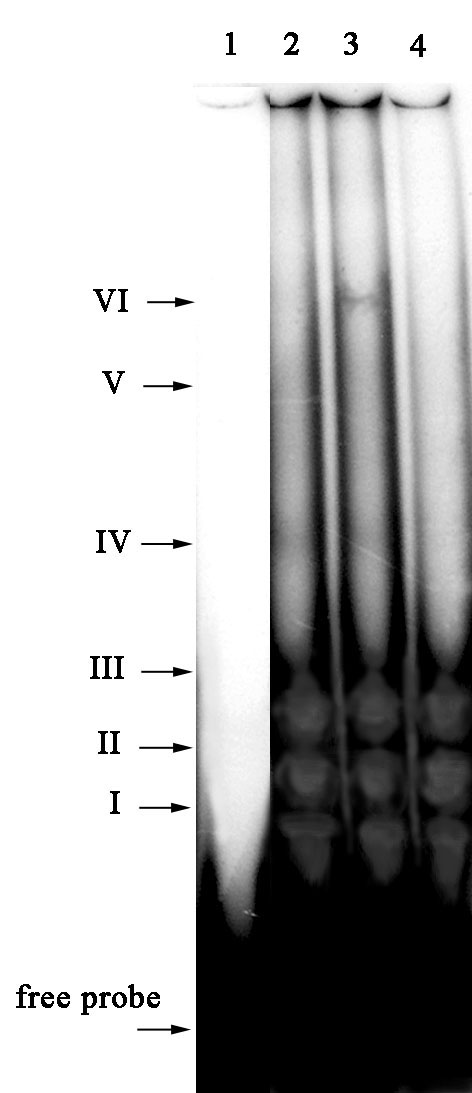
Surprisingly, the effects of the glutathione redox system on DNA binding activity of nuclear topo I were quite different when the oligonucleotide containing core sequence of the atp9 gene promoter was used as a DNA probe (Fig. 2, I-VI). A supplementary protein-DNA complex appeared upon addition of GSH into retardation medium (Fig. 2, VI). In our previous report (MNL 77:37-38, 2003), we showed the same phenomenon of stimulation of DNA binding activity of mitochondrial topo I in the presence of GSH. It should be noted that when using the atp9 probe, the addition of GSSG, on the contrary, caused an inhibitory effect on DNA binding activity of nuclear topo I, consisting of the complete loss of slower migrating retardation complexes (Fig. 2, IV-VI).
Consequently, such a universal physiological redox modulator as glutathione has a pronounced influence on DNA binding activity of DNA topoisomerase I of nuclear origin. In our opinion, the data received speak well for the existence of a redox regulation mechanism involving not only modulation of the activity of transcriptional factors, but also a topo I redox modulation mechanism based on modification of cysteine residues in the protein molecule of nuclear and mitochondrial DNA topoisomerase. Future experiments on site-directed mutagenesis will allow discovery of the possible role of individual cysteine residues in redox modulation of DNA binding and catalytic activity of topo I. Additional experiments are needed to elucidate the cause of the demonstrated differences in effects of the glutathione redox system on nuclear topo I DNA binding activity when using different DNA substrates.
Financial support from the Russian Foundation for Basic Research (Grant No. 01-04-48162) is acknowledged.
Figure 1. The effect of reduced and oxidized glutathione on DNA binding activity of nuclear topoisomerase I with the p53 gene promoter fragment. 1) control (-topo I), 2) topo I, 3) topo I + GSH, 4) topo I + GSSG.
Figure 2. The effect of reduced and oxidized glutathione on DNA binding activity of nuclear topoisomerase I with the atp9 gene promoter fragment. 1) control (-topo I), 2) topo I, 3) topo I + GSH, 4) topo I + GSSG.
{kind=link}
{kind=link}