Córdoba, Argentina
Universidad Nacional de Córdoba
Preliminary studies on combining ability effects for canopy temperature in corn (Zea mays L.)
--Biasutti, CA, Alemanno, GA
Canopy temperature has been indicated as a trait to discriminate among drought tolerant and non-tolerant maize genotypes. There are no reports on the gene action responsible for canopy temperature in corn. The objective of this work was to study the gene action for canopy temperature in maize and its impact on grain yield.
Ninety-three breeding lines were screened during two consecutive years for canopy temperature (CT). A hand-held infrared thermometer was employed to take canopy temperature. Temperature was recorded in breeding trials from 10 plants of each of 2 replications of each line in each year. Five measures were taken form the same plant during the vegetative phase in each line. Temperature was recorded around noon (11 – 13 hs) on sunny cloudless days. Medium values for each line were utilized to classify them as high (HCT) or low (LCT) canopy temperature lines. No other agronomic attributes were considered when the lines were selected for CT. The inbreds 6A, 11 and 86C were classified as LCT and inbreds 83A, 48B and 70A as HCT. All inbreds were derived from the same population by the pedigree selection method.
During the summer of 2001/2002 fifteen single cross hybrids among the six inbred lines selected for high or low canopy temperature were produced by the diallel-mating scheme.
In the summer of 2002/2003, a field experiment with 23 entries (15 single crosses, 6 parental inbreds and 2 checks) was conducted at the Experimental Station of Facultad de Ciencias Agropecuarias de Córdoba, in Córdoba province. The design was in randomized complete block, with three replications in plots 7 m long and spaced 0.7 apart, with 46.6 plants per plot. Canopy temperature of all entries was recorded in the same way as described above. Grain yield was recorded on a plot basis and corrected at 14% grain moisture.
A preliminary analysis of variance was made for CT and GY. The general combining ability (GCA) and specific combining ability (SCA) information were obtained by using the method 2 model I analysis of Griffing (Austr. J. Biol. Sci. 9:463–493, 1956), for the variables canopy temperature (CT) and grain yield (GY).
The inbred and hybrid means for canopy temperature and grain yield are in Table 1. The analysis of variance of the diallel crosses indicated significance for GCA and SCA for the two traits analyzed (Table 2). The canopy temperature and yield mean squares for SCA were larger than for GCA. The proportion of GCA in relation to SCA can be calculated by GCA/(GCA+SCA) (Baker, Crop Sci. 18:553–536, 1978). The values of this relation for CT and yield were 26 and 36% respectively, indicating that nonadditive genetic effects are very important. It is known that, in maize, nonadditive effects are generally more important for yield than for other traits, as indicated by Hallauer and Miranda (Quantitative analysis in maize breeding, Iowa State Univ. Press, Ames, p. 468, 1988). The greater expression of nonadditive effects for yield must be partially due to the presence of deleterious genes that cause inbreeding depression.
Low CT Inbred lines showed negative GCA effects, whereas high CT lines showed positive GCA effects (Table 3). Inbred 83A (HCT) had the largest GCA estimate for CT. Inbred 48B had the lesser GCA estimate for CT and a large negative GCA estimate for yield.
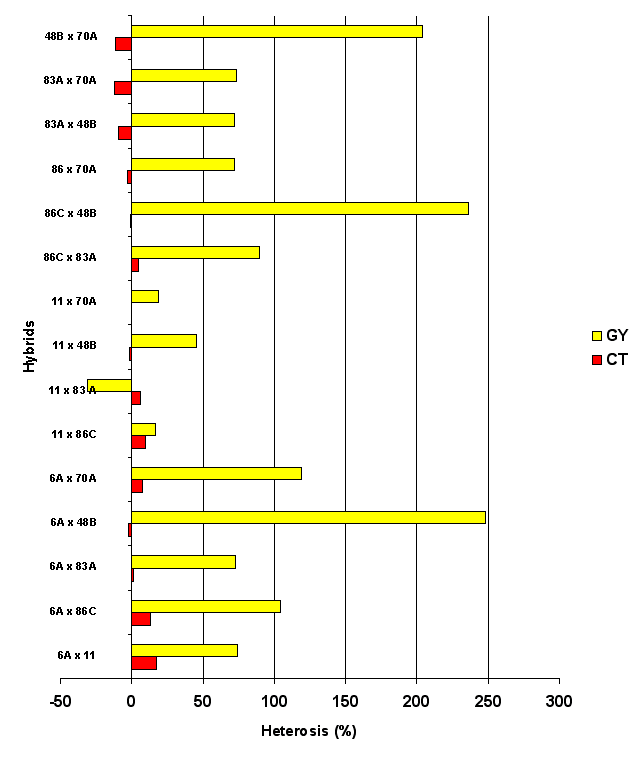
The SCA estimates obtained by the Griffing (1956) method are equivalent to the specific heterosis obtained by the Gardner and Eberhart (Biometrics 22:439–452, 1966) analysis. Crosses with major positive heterosis for grain yield showed little or negative heterosis for canopy temperature (Figure 1). According to the data analyzed we can conclude that there is variability for CT among the inbred lines and so it is possible to select for low or high canopy temperature in breeding lines. High yield heterosis was found in LCT × HCT crosses. Further research is needed, with data from different environments, to confirm or negate these conclusions.
Table 1. Means of 6 maize inbred lines and 15 diallel crosses evaluated for canopy temperature and grain yield.
| Genotypes | Canopy Temperature °C |
Grain yield q/ha |
| 6A | 23.46 | 13.69 |
| 11 | 23.73 | 47.45 |
| 86C | 24.70 | 32.74 |
| 83A | 28.03 | 43.81 |
| 48B | 28.33 | 10.71 |
| 70A | 28.27 | 29.76 |
| 6A × 11 | 27.62 | 53.19 |
| 6A × 86C | 27.32 | 47.41 |
| 6A × 83A | 26.10 | 49.75 |
| 6A × 48B | 25.26 | 42.44 |
| 6A × 70A | 27.85 | 47.66 |
| 11 × 86C | 26.54 | 46.67 |
| 11 × 83A | 27.54 | 31.54 |
| 11 × 48B | 25.52 | 42.22 |
| 11 × 70A | 25.86 | 45.82 |
| 86C × 83A | 27.64 | 72.62 |
| 86C × 48B | 26.28 | 73.03 |
| 86C × 70A | 25.70 | 53.66 |
| 83A × 48B | 25.56 | 46.87 |
| 83A × 70A | 24.80 | 63.91 |
| 48B × 70A | 25.10 | 61.48 |
| LSD | 1.35 | 13.24 |
Table 2. Analysis of variance of 15 diallel crosses among six maize inbred lines for canopy temperature (CT) and grain yield (GY), according to Method 2 of Griffing (1956).
| Source | d.f. | Mean squares* | |
| CT | GY | ||
| Genotypes | 20 | 6.48 | 764.63 |
| GCA | 5 | 2.89 | 487.81 |
| SCA | 15 | 7.68 | 856.90 |
| Error | 40 | 0.67 | 64.40 |
*F tests were significant in all instances at P<0.01
Table 3. Estimates of general and specific combining ability effects for inbred lines and for hybrids, respectively, for canopy temperature and grain yield in maize.
| Genotypes | Canopy Temperature | Grain Yield |
| 6A | -0.333 | -6.372 |
| 11 | -0.399 | -0.558 |
| 86C | -0.107 | 5.007 |
| 83A | 0.495 | 4.187 |
| 48B | 0.080 | -3.918 |
| 70A | 0.264 | 1.655 |
| 6A x 11 | 2.104 | 14.57 |
| 6A x 86C | 1.512 | 3.230 |
| 6A x 83A | -0.310 | 6.390 |
| 6A x 48B | -0.735 | 7.186 |
| 6A x 70A | 1.671 | 6.833 |
| 11 x 86C | 0.798 | -3.323 |
| 11 x 83A | 1.196 | -17.633 |
| 11 x 48B | -0.408 | 1.153 |
| 11 x 70A | -0.252 | -0.820 |
| 86C x 83A | 1.003 | 17.880 |
| 86C x 48B | 0.058 | 26.396 |
| 86C x 70A | -0.705 | 1.453 |
| 83A x 48B | -1.263 | 1.056 |
| 83A x 70A | -2.207 | 12.523 |
| 48B x 70A | -1.492 | 18.199 |
Figure 1. Heterosis for canopy temperature (CT) and grain yield (GY) in 15 maize hybrids.
{kind=link}