Five cycles (periods) have determined breeding and growing of hybrid maize in Yugoslavia. Each period has been characterized by the introduction of new higher yielding hybrids. The increase of genetic yield potential of selected hybrids over these cycles amounted to 1,094 t, or approximately 100 kg per year (Ivanovic et al, Symposium of Maize Breeding, Production and Utilization, pp.3-17, Belgrade, Sept. 1995).
The first cycle of selection is represented by the first local double cross hybrids, replacing the formerly grown American double cross hybrids. They were introduced into broader production at the end of the 50's and the beginning of the 60's. During the 60's, remarkable progress was achieved during the second cycle, when the first single cross hybrids were introduced. They increased genetic potential and uniformity, which led to their predominant use and production during the 70's. The hybrids of the third cycle were introduced at the end of the 70's. These were the new single cross hybrids, which had increased genetic yield potential 10-15% and were more tolerant to high density cropping. Fourth cycle hybrids were introduced at the end of the 80's, and had increased drought tolerance and increased tolerance to stalk lodging. The newly developed hybrids respond better to a higher level of cropping practices and achieve maximum genetic yield potential at greater plant densities (Drinic et al, TOSS 12, pp.1-8, 2001).
Knowledge of the genetic diversity and relationships among maize hybrids is important for planning breeding strategies, hybrid identification, and germplasm identification. The genetic similarity of two genotypes could be estimated indirectly from pedigree information or directly by molecular markers (isozymes, protein, DNA markers). The utility of protein markers to characterize maize hybrids, validate pedigrees, and show association among hybrids was evaluated using a set of 30 ZP maize hybrids from 5 different selection periods (6 hybrids from each cycle).
The proteins were isolated from embryos of hybrids from different selection periods and separated by PAGE (Polyacrylamide Gel Electrophoresis) according to Leammli (Leammli V.K., Nature 227:680-685, 1970). The analysis of embryo salt soluble proteins shows that all genotypes studied have a specific protein pattern. Both quantitative and qualitative differences between protein fraction were determined, and indices of similarity were calculated.
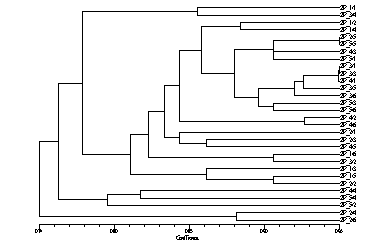
The UPGMA clustering method was used for hierarchical clustering and the necessary computations were performed using the NTSYS-pc program (Rohlf, New York Exeter Publ, 1989). The protein based dendrogram, consisting of three major groups, is presented in Figure 1. The first group is further divided into seven subclusters, assigned from A to F. Twenty-five hybrids were distributed within one large and six small subgroups The main subcluster C includes 13 ZP hybrids, mainly from the third, fourth and fifth cycles of selection. It was comprised of hybrids derived from or related to BSSS germplasm. The hybrids that have Wf9 and Oh43 lines as one parent grouped together in subcluster G and E, respectively. The subcluster A, consisting of two hybrids, one from the first and one from the third cycle of selection, was loosely aggregated with a large group of hybrids.
Figure 1. Dendrogram of 30 maize ZP hybrids based on protein markers
The five remaining hybrids with one parent from or related to Lancaster germplasm form two separate groups. Three hybrids, two from the fifth and one from the fourth cycle of selection, form the second group. The third group consists of two hybrids belonging to the second cycle of selection with the common parent line C103.
Grouping of 30 ZP hybrids from different cycles of selection generally agreed with the pedigrees, but some discrepancies in forming subclusters within major groups were noted. One discrepancy was that two double cross hybrids, ZP1/4 and ZP1/3 from the first cycle of selection, which had three of four parental inbred lines in common, were in different subclusters. Also, hybrid ZP2/5, instead of being in the first group, subcluster C, has to be in the third group due to its orgin.
In summary, our study provides information on the molecular basis of
polymorphism detected by protein markers in 30 ZP maize hybrids. It demonstrates
that protein markers can identify and validate genetic relationships among
maize hybrids.
Return to the MNL 77 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}