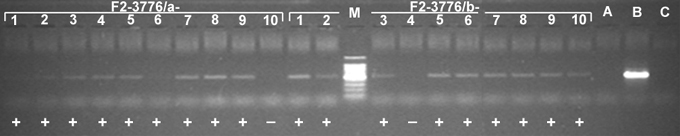
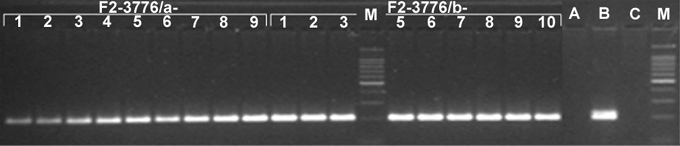
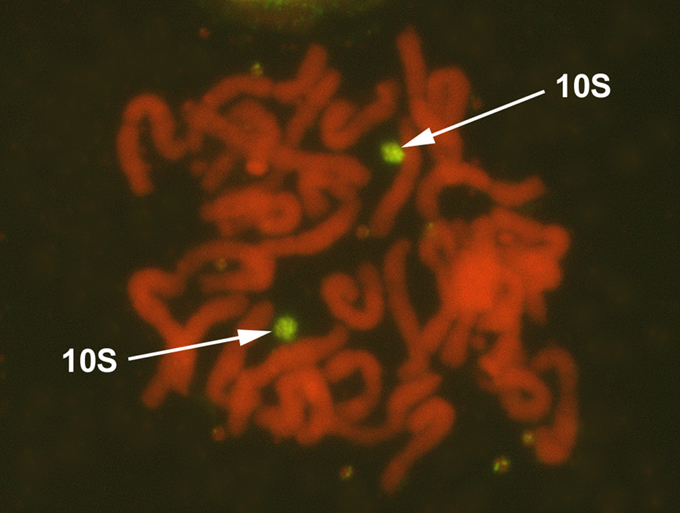
Plant �F1-0289-1� retained the maize chromosome 10, in addition to all oat chromosomes, based on a positive test for the maize-specific LTR-type retrotransposon Grande 1 and maize chromosome 10 SSR markers. The plant was allowed to self-pollinate, and four maize-positive panicles were found to set seeds (Table 1). Genomic DNA samples of ten F2 offspring from each maize-positive panicle of the F1-0289-1 plant were assayed for presence versus absence of the maize-specific LTR-type retrotransposon Grande 1 (Figure 1). PCR using Grande 1-specific primers detected maize chromatin in nine offspring plants of F1-0289-1/a (designated F2-3776/a-1 to F2-3776/a-9; F2-3776/a-10 was negative), nine offspring plants of F1-0289-1/b (designated F2-3776/b-1 to F2-3776/b-10; F2-3776/b-4 was negative), no offspring plant of F1-0289-1/c (all F2-3776/c-1 to F2-3776/c-10 were negative), and ten offspring plants of F1-0289-1/d (designated F2-3776/d-1 to F2-3776/d-10). We verified the maize chromosome identities by assaying the corresponding DNA samples of the maize-positive F2 offspring plants with two sets of SSRs genetically mapped to maize chromosome 10 from the MaizeDB We selected the markers: p-phi041, p-phi117, and p-umc1293 from bin 10.00, and p-umc1249, p-umc1196, p-umc1176 and p-umc1084 from bin 10.07. Since most of the maize-positive plants showed consistent presence of the three markers from the bin 10.00, but absence of the four markers from the bin 10.07, we conclude that the maize chromosome present in each of these offspring is likely a telocentric derivative of the chromosome 10 retained in the original plant �F1-0289-1�. GISH analysis on root-tip cells using labeled genomic DNA from maize revealed the disomic status and the telocentric nature of the added chromosomes in most of the plants analyzed by PCR (Figure 3). The line was given the designation OMAdt10.20S in accordance with the proposed nomenclature for oat-maize chromosome addition lines (Kynast et al. 2001: Maize Genetics Coop. Newsl. 75, 54-55).
Since there are more seeds to be analyzed for the possible telosomic addition of the long arm of chromosome 10, we will test all seeds with larger numbers of chromosome arm-specific SSRs. At present we are propagating the identified disomic telocentric F2 offspring plants to test for chromosome stability and transmission to F3 offspring. Once more seed are produced, the new addition lines will be available upon request to the scientific community.
This material is based upon work supported by the National Science Foundation under Grant No. 0110134.
Table 1. Seed set of the first fertile oat-maize chromosome 10 addition
F1 (Sun II Seneca 60) 0289-1.
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Figure 1. Selected examples of oat offspring possessing maize-chromatin shown by PCR with Grande 1 primers. The presence of the 500 bp band (+) indicates presence of maize chromatin in the DNA extract of the corresponding F2 plant; M=Molecular Weight Marker, A=Sun II, B=Seneca 60, C=No-DNA Control.
Figure 2. Telocentric maize chromosome identification with the short arm-specific SSR, p-phi041 (bin10.00). The presence of the 200 bp band is diagnostic for the short arm of maize chromosome 10 as shown in DNA extracts of the corresponding F2 plants; M=Molecular Weight Marker, A=Sun II, B=Seneca 60, C=No-DNA Control.
Figure
3. GISH of fluorophore-labeled genomic maize DNA to root meristem cells
of plant F2-3776/b-6. Arrows point to the two telocentric maize chromosomes
(green), which are formed from the short arm of chromosome 10 (smallest
maize chromosome). Oat chromosomes (red) are counterstained with propidium
iodide.
Return to the MNL 77 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}