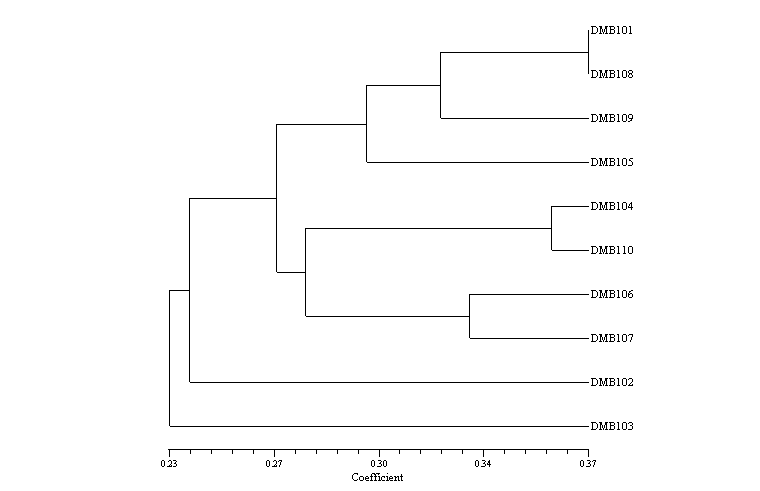
DMB 107 and DMB 108 (derived from A-64 source population) clustered together. Also, the similarity of DMB 110 and DMB 104, which shared a common source population, was implied. However, many discrepancies were noticed in the general clustering patterns (on the basis of morphological and SSR markers individually or in combination). This could be possibly due to a limited number of traits, low SSR markers, multiple underlying genes for these traits and scoring errors. The broad base of the source population, as well as over-representation of a particular source population (for example, five of the ten genotypes were derived from A-64), might be the reason for the low level of conformity between genetic relationships vis a vispedigree information. It is not uncommon to expect inconsistency between SSR grouping and genetic background of corn inbreds, and this may be attributed to many factors (Yu et.al., Maydica 46:133-139, 2001).
Figure 1. Dendrogram depicting genetic relationships among selected inbred based on morphological and SSR data
Superior heterotic single cross combinations in comparison to PEHM-2 (the check with comparable maturity) were identified (DMB 101 x DMB 109, DMB 102 x DMB 103, DMB 102 x DMB 110, DMB 103 x DMB 104 and DMB 104 x DMB 110) on the basis of per se performance (Table 1). Of the five elite hybrid combinations identified over two locations, three crosses (highlighted) involved the parental line belonging to different clusters generated by the combined data of morphological and molecular markers. Hence, the clustering pattern can be taken as a general indicator for choosing potential heterotic combinations. The relationship between hybrid relatedness and/or pedigree information, in terms of heterosis performance, can be explained and interpreted (Smith and Smith, MNL 63:86-87, 1989; Smith et.al., Maydica 45:235-241, 2000)
The fact that all parental inbred lines involved in these crosses (except DMB 110) were derived from the same source population, A-64 (Table 2), gave further credence to the broad base of the source population.
Table 1. Particulars and performance of promising maize single cross
hybrids
|
|
|||||||
|
|
Tasseling |
Silking |
|||||
| S. No | Cross |
|
|
|
|
|
|
| 1. | DMB 103 X 104 |
|
|
|
|
|
|
| 2. | DMB 101 X 109 |
|
|
|
|
|
|
| 3. | DMB 102 X 103 |
|
|
|
|
|
|
| 4. | DMB 104 X 110 |
|
|
|
|
|
|
| 5. | DMB 102 X 110 |
|
|
|
|
|
|
Table 2. Particulars of maize inbred lines involved in elite single
cross hybrids
| Inbred No. | Pedigree | Source Population |
| DMB 101 | IPA 3-6-10-3-1-1-1-2-1-# | A-64 |
| DMB 102 | IPA 3-f (-1) | A-64 |
| DMB 103 | IPA 3-f (-2) | A-64 |
| DMB 104 | IPA 1-f-16-2-#-f-1 | A-64 |
| DMB 109 | TCA 22-3-1-1-1-f-#-f-1 | A-64 |
| DMB 110 | SC 7-2-1-1-7-1-1-1-1 | Derived from MDR-1 X A-64 |
In the present investigation aimed at characterization of selected maize
inbreds, SSR markers were instrumental in finer discrimination of inbred
lines, as well as more precise analysis of homozygosity. Though differentiation
of maize lines on the basis of morphological traits is presently contemplated,
for finer discrimination between the parental lines, molecular markers
like SSRs will be used. Some discrepancy in clustering could be accounted
for by various factors. Molecular markers can serve as an invaluable aid
for a variety of applications in maize breeding (Dudley et.al., Crop Sci.
31:718-723, 1991).
Return to the MNL 77 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}