Three-day-old etiolated shoots of maize (Zea mays L. cv. Rossiyskaya) germinated on moist paper at 26 C were used in this work. Shoots were cold-stressed at 0 C for 1 h or were hardened at 8 C for 7 days. After cold treatment, mitochondria were isolated from seedlings shoots by differential centrifugation (Pobezhimova et al., J. Therm. Biol. 21:283-288 1996) and their energetic activity was studied. The mitochondria isolated were resuspended in the following medium: 40 mM MOPS-KOH buffer (pH 7.4), 300 mM sucrose, 10 mM KCl, 5 mM EDTA, 1 mM MgCl2. The activity of mitochondria was recorded polarographically at 27 C using a platinum electrode of a closed type in a 1.4 ml volume cell. The reaction mixture contained 125 mM KCl, 18 mM KH2PO4, 1 mM MgCl2 and 5 mM EDTA, pH 7.4. During NADH oxidation to incubation media, Ca2+ was added (0.06 mM CaCl2) (Moller et al., Biochem. J. 194:487-495 1981). The concentration of mitochondrial protein was analysed by the Lowry method (Lowry et al., J. Biol. Chem. 1059:265-275 1951). Polarograms were used to calculate the rates of phosphorylative respiration (state 3), non-phosphorylative respiration (state 4), respiratory control by Chance-Williams and the ADP:O ratio (Estabrook, Methods Enzymol. 10:41-47 1967). All the experiments were performed on 3-6 separate mitochondrial preparations.
It was found that maize mitochondria that used malate as oxidation substrate were the most sensitive to cold shock and cold hardening, as compared with succinate and NADH oxidizing mitochondria. Cold shock and hardening caused about a 40% increase of state 4 respiration, a 26% decrease of RC coefficient, and a 10% decrease of ADP:O ratio. Such changes in mitochondrial energetic activity are typical for a low-energetic mitochondrial state.
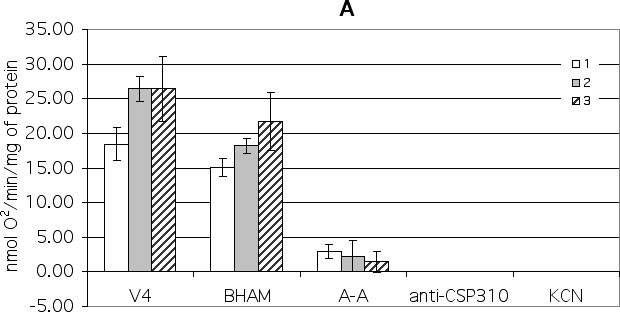
To study the participation of cytochrome and alternative electron transport pathways, the effects of sequential additions of benzhydroxamic acid (BHAM) (an inhibitor of AO) and antimycin A (an inhibitor of complex III electron transport) on the activity of isolated maize mitochondria were used. It was shown that mitochondria isolated from control, stressed and hardened seedling shoots differ in their reaction when these inhibitors were added. Indeed, the addition of BHAM to control mitochondria oxidising malate in state 4 caused a slight decrease (18%) in the rate of oxygen uptake (Fig. 1A). A subsequent addition of antimycin A inhibited the rate of oxygen consumption up to 14%. The remaining oxygen consumption was fully inhibited by the addition ofanti-CSP310 antiserum (Fig 1A). So, in non-stressed maize mitochondria the following pathways are functional: cytochrome pathway (66%), AO (18%), and CSP 310 (16%). Cold shock caused the decrease of the main cytochrome pathway (60%), a significant increase of the AO pathway (31%), and a slight decrease of the CSP 310-pathway (8%) (Fig. 1A). Hardening caused an increase of the main cytochrome pathway (77%), did not influence the AO pathway (18%), and caused a slight decrease of the CSP 310-pathway (5%) (Fig. 1A).
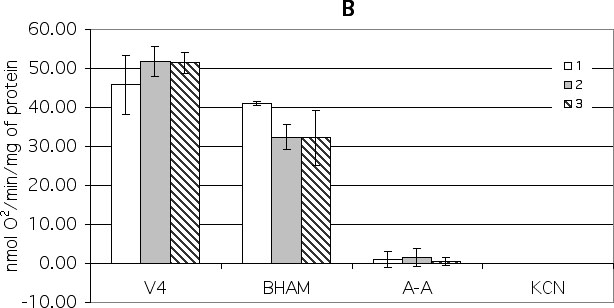
On the other hand, an addition of BHAM to control succinate-oxidizing state 4 maize mitochondria resulted in a 10% inhibition of oxygen uptake (Fig. 1B). A subsequent addition of antimycin A fully inhibited mitochondrial respiration. So, in succinate-oxidizing maize mitochondria, both the main cytochrome (90%) and AO pathway (10%) function. Cold stress caused a decrease of the cytochrome pathway (60%) and an increase of the AO pathway (38%) (Fig. 1B). Cold hardening caused the decrease of cytochrome pathway (62%) and an increase of AO pathway (38%) too (Fig. 1B). In all cases, the sequential addition of BHAM and antimycin A to succinate-oxidizing mitochondria fully inhibits the oxygen uptake.
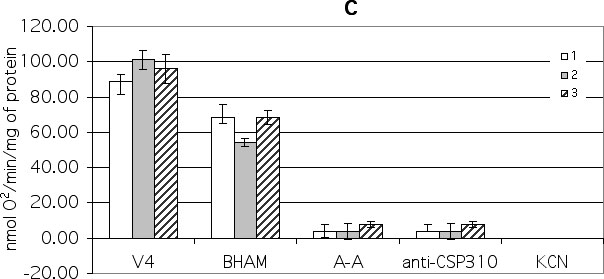
The addition of BHAM to NADH-oxidizing state 4 control maize mitochondria resulted in a 23% inhibition of oxygen uptake, and addition of antimycin caused about a 73% inhibition of mitochondrial respiration (Fig. 1C). The remaining oxygen uptake after BHAM and antimycin A addition was about 5%, and was not affected by anti-CSP 310 antiserum addition. It is possible that this oxygen uptake depends on the presence of the external antimycin A-insensitive NADH-cytochrome c reductase (Soole et al., Physiol. Plantarum 78:205-210 1990). This residual respiration was fully inhibited by cyanide (Fig. 1C). The addition of BHAM to cold-stressed maize mitochondria caused a 46% inhibition of oxygen uptake, and subsequent antimycin A addition inhibits mitochondrial respiration up to 4% (Fig. 1C). Cold hardening caused the decrease of the main cytochrome pathway (63%), a slight increase in the AO pathway (29%), and an increase in the antimycin A and anti-CSP310 resistant pathways (8%) (Fig. 1C). So, in NADH-oxidizing maize mitochondria the main cytochrome pathway, the AO pathway and the "external" antimycin A-resistant pathways are present. The main cytochrome pathway is the most active in maize mitochondria (73%), but it decreases during short-term cold stress and cold hardening.
So, based on the data obtained we can detect the presence of different electron transport pathways in maize mitochondria. The combination of different NAD(P)H dehydrogenases, AOX and CSP 310 should make plant mitochondria more tolerant to different stress factors.
Acknowledgements: The work has been performed, in part, with the support of the Russian Foundation of Basic Research (projects 00-04-48093 and 02-04-06096) and the Siberian Division of Russian Academy of Sciences Youth Grant (project 78).
Figure 1. The effect of different respiratory chain inhibitors on state 4 oxygen consumption by maize mitochondria isolated from control (1), stress (2) and cold-hardened (3) shoots. The inhibitors were added in the sequence shown reading left to right. Their final concentrations were: - BHAM (1 mM), antimycin A (A-A) (20 mkM), KCN (0.4 mM) and CSP 310 antiserum (1 mg/mL).
A. 10 mM malate in the presence of 10 mM glutamate was used as a substrate of oxidation.
B. 8 mM succinate in the presence of 5 mM glutamate and 3 mkM rotenone was used as a substrate of oxidation.
C.
1 mM NADH in the presence of 3 mkM rotenone was used as a substrate of
oxidation.
Return to the MNL 77 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}