The role of smHSPs during oxidative and water stress remains unclear, although the study of oxidative and water stress protection mechanisms is an important task. Agricultural plants are often exposed to drought in natural conditions. Oxidative stress is known to accompany many other stress impacts (Davies and Goldberg, J. Biol. Chem. 262:8220-8226, 1987).
Wehmeyer and Vierling (Plant Physiol. 122:1099-1108, 2000) showed that the ability to overcome dehydration correlates with the ability to synthesize smHSPs in mutant drought-resistant Arabidopsis thaliana seeds. The same regularity was observed in desiccation-intolerant Craterostigma plantagineum callus tissue (Alamillo et al., Plant Mol. Biol. 29:1093-1099, 1995). Nevertheless, up to now smHSP induction in response to water stress has not been directly proven, apart from the work by Almoguera and Jordano (Plant Mol. Biol. 19:781-792, 1992). They demonstrated smHSP accumulation compared to control in sunflower seedlings after 4 and 24 hours of osmotic stress created by mannitol in the vegetation solution.
Oxidative stress was shown to induce smHSP accumulation in rice chloroplasts (Lee et al., Gene 245:283-290, 2000). The key role of smHSPs in the protection of photosystem II of chloroplasts from oxidative stress was proven (Downs et al., J. Plant Biol. 155, 1999). However, it is still unknown whether smHSPs are accumulated elsewhere in the cell in response to oxidative stress.
The objective of our work was to check whether smHSP accumulation takes place in maize seedlings under long-term and short-term dehydration created by different methods. We also intended to determine whether smHSPs are accumulated elsewhere in the cell, apart from chloroplasts, in response to oxidative stress.
We analyze total cell proteins from etiolated maize seedlings subjected to oxidative and water stress by SDS-PAGE and immunoblotting using antibodies developed for smHSP-specific C-terminal a-crystallin domain (Heckathorn et al., Plant Physiol. 116:439-444, 1998). Simultaneously we monitored water stress. We used three-day-old etiolated maize seedlings grown at 27 C. To provide oxidative stress we submerged the seedlings in the solution containing 100 mM of methyl viologen, a herbicide provoking free oxygen radical production in the cell, for 3 hours. Dehydration was created by two methods. Three-day-old seedlings were left in thermostat for 24, 48 or 72 hours without water. Then the distinct groups of seedlings were incubated in solutions containing 38, 64 or 88 mM of polyethyleneglycol (PEG), which corresponded to 300, 500 and 700 mOsm respectively. To determine moisture content in the seedlings they were dried to dry weight at 80 C for 20 hours with the weight measured before and after drying. Total proteins were extracted from seedlings as described elsewhere (Borovskii et al., J. Plant Physiol. 156:797-800, 2000). Proteins were subjected to SDS-PAGE (14% of acrylamide) using a mini-Protean II cell (Bio-Rad, USA) according to the manufacturer�s instructions. Western blot and immunodetection were carried out as was described previously (Timmons and Dunbar, Methods Enzymol. 182:679-688, 1991). Antibodies to the a-crystallin conserved C-terminal domain, common to all eukaryotic smHSPs and to the a-crystallin proteins of the vertebrate eye lens, kindly provided by Dr. Craig A. Downs, were used for detection of smHSPs (Heckathorn et al., Plant Physiol. 116:439-444, 1998).
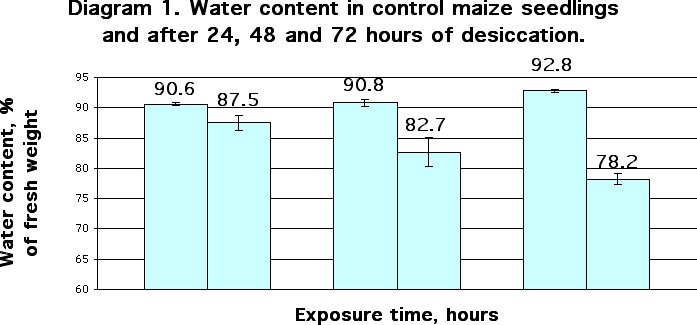
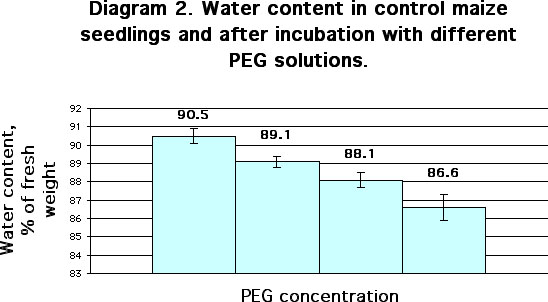
Significant dehydration of seedlings was shown to take place after PEG treatment and desiccation. Thus, after 24 hours of drought stress water content in the seedlings decreased from 90.6 % (control) to 87.5%. After 48 and 72 hours, water content amounted to 82.7% and 78.2%, respectively (Diagram 1). After incubation of the seedlings with PEG solutions of 300, 500 and 700 mOsm water content decreased from 90.5% (control) to 89.1, 88.1 and 86.6% respectively (Diagram 2).
Immunochemical analysis of total cell protein produced the following results. A small amount of smHSPs were present in the seedlings in the conditions of optimal moisture content, that is, in the control (Fig. 1, Fig. 2). This agrees with our previous results (Korotaeva et al., MNL 75:24-25, 2001). SmHSP accumulation among total cell proteins positively correlated with the duration and intensity of water stress. Thus, well pronounced accumulation of smHSPs with Mr 19 and 22 kD was observed, as compared to the control, after 48 and 72 hours of desiccation, and after incubation with PEG solution of maximal osmotic pressure (700 mOsm) (Fig. 1). However, we did not manage to identify smHSP accumulation among total cell protein with seedlings treated by methyl viologen (Fig. 2). This agrees with the results of Banzet and co-authors (The Plant J. 13:519-527, 1998), shown in tomato suspension cultures, that smHSPs do not accumulate in the cells as a response to treatment by O2 (o-) generating agents methyl viologen and digitonin, though smHSP accumulation happens with H2O2 treatment. Nevertheless, other researchers showed that pea leaf treatment by methyl viologen still brings about the consequences pointing to the presence of oxidative stress. This was witnessed by the decrease (compared to control) in concentration of some antioxidant enzymes, and the accumulation of oxidized proteins and catalytic Fe in pea leaf parenchyma (Iturbe-Ormaetxe et al., Plant Physiol. 116:173-181, 1998). Therefore, the consequences resulting from methyl viologen treatment do not induce smHSP accumulation. It is still unknown what kind of oxidative stress consequences lead to smHSP accumulation in the cell.
This work was supported by the Russian Fund of Basic Research (projects 02-04-06022 and 02-04-48599).
Figure 1. Accumulation of smHSPs in maize seedlings during desiccation and PEG treatment. Seedlings were grown at 27 C in optimal water conditions. One group of three-day-old seedlings was kept without water for 24, 48 or 72 hours. Others were cut and incubated 3 hours with PEG solution of 300, 500 or 700 mOsm. Seedlings kept without treatment were used as control. Total protein fraction was divided by SDS-PAGE (14% of acrylamide), and proteins were transferred from gel to nitrocellulose membrane, which was incubated with primary antibodies (1:500 dilution), elaborated to the a-crystallin domain (Heckathorn et al., Plant Physiol. 116:439-444, 1998). Molecular masses of smHSPs are given on the left.
Figure 2. SmHSPs of maize seedlings at control conditions and after methyl viologen treatment. Three-day-old seedlings were cut and incubated 3 hours either with 100 mM of methyl viologen dissolved in 0.01% of Tween 100 (MV) to induce oxidative stress, or in 0.01% of Tween 100 for control. Total protein fraction was divided by SDS-PAGE (14% of acrylamide), and proteins were transferred from gels to nitrocellulose membrane, which was incubated with primary antibodies (1:500 dilution), elaborated to the a-crystallin domain. Molecular masses of smHSPs are given on the left.
Seedlings were grown at 27 C in optimal water conditions. One group of three-day-old seedlings was kept without water at 27 C for 24, 48 or 72 hours (columns 2, 4, 6, respectively). Other seedlings were kept 24, 48 or 72 hours in optimal water conditions at the same temperature (columns 1, 3, 5, respectively). After that we carried out detection of water content in seedlings. Each replication contained three measurements of water content in groups of 40-50 shoots. Mean ± S.D (n=3; P=0.95).
Seedlings were grown at 27 C in optimal water conditions. Three-day-old
seedlings were cut and divided into four groups of 30-40 shoots. Each group
was distinctly incubated 3 hours with PEG solutions of 300 mOsm (column
2), 500 (column 3) or 700 mOsm (column 4). Seedlings incubated with distilled
water were used as control (column 1). After that, detection of water content
was carried out. Each replication contained three measurements in groups
of 30-40 shoots. Mean ± S.D (n=3; P=0.95).
Return to the MNL 77 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}