The 22 kDa zein (z1C) gene subfamily has two different genomic locations. The z1C gene cluster contains most family members except for one, and is physically linked to the php200725 RFLP marker on maize chromosome 4S (~55 kb apart). The single copy member of the 22 kDa zein genes (asz22;16) has translocated to a different position on the same chromosome arm. Using a single-nucleotide-polymorphism (SNP) in the 22 kDa zein gene coding region between two different inbred lines, BSSS53 and Mo17, we were able to map this gene member by a (Mo17xBSSS53)xMo17 backcross mapping population. A primer pair: (5� primer: TCACTTGCTCCTAGTGGCAG; 3� primer: TAGATGAAAGTAGTTGTAGGTAGA), which will only give a PCR product from azs22;16 of BSSS53 but not that of Mo17, was used for scoring the recombinant events from the population. The same population was also used to map the php200725 and cdo520 loci relative to the location of the zein genes. Out of 107 samples, we found 21 recombinant events between php200725 and azs22;16, which gave a genetic distance of 19.6 cM. We also found four recombination events between php200725 and cdo520, and 17 recombination events between cdo520 and azs22;16. Therefore cdo520 was placed in between php200725 and azs22;16, with 3.7 cM genetic distance to php200725. Sequence homology comparison indicated that azs22;16 is the normal allele of floury2.
The 19 kDa zeins consist of three gene subfamilies (Song, R. and Messing, J, Plant Physiology 130:1626-1635, 2002). According to BAC clone analysis, the two larger subfamilies, z1A and z1B, both have two different genomic locations. The smallest subfamily, z1D, only comprises one genomic location. Using a z1D subfamily-specific probe, we found that the z1D subfamily is located on maize chromosome 1 by using oat-maize addition lines (data not shown). Previously, Woo, YM et al. (Plant Cell 13:2297-2317, 2001) mapped the two z1D genes, az19D1 and az19D2, to positions 123.3 and 122.4, consistent with our results. The 0.9 cM genetic distance between the two z1D genes reflects a ~200 kb physical distance according to our sequence analysis of the z1D gene subfamily (Song, R. and Messing, J, Plant Physiology 130:1626-1635, 2002).
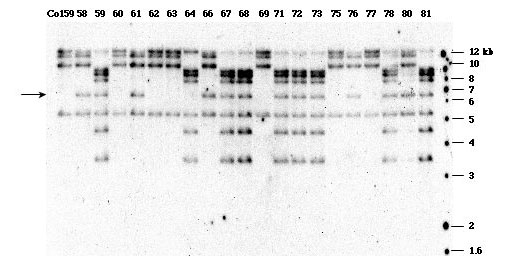
The other two 19 kDa zein gene subfamilies both comprise two different genomic locations. Each genomic location contains gene clusters with high sequence homology indicating recent gene amplifications within short physical intervals. The subfamily-specific probes that were used to isolate the BAC clones were not sufficient to distinguish the two locations. Therefore instead of mapping the two locations by location-specific probes, we decided to map them based on their co-segregation pattern using a subfamily-specific probe. The subfamily-specific probe will detect all the bands from both genomic locations on a Southern blot, but only those bands belonging to the same location will co-segregate. We used the z1A and z1B-specific probes that were previously described (Song, R. and Messing, J, Plant Physiology 130:1626-1635, 2002) to check for sequence polymorphism of the parental lines of two maize recombinant inbred lines (Burr, B et al., Genetics 118:519-526, 1988). Based on a maximum of polymorphism detected by Southern blot analysis (number of bands and their polymorphism), we chose the Tx303xCo159 population cut with EcoRI for z1A subfamily. One band does not show polymorphism between the two parental lines. However, other bands could be sorted into two co-segregating groups (Fig. 1). One group contains most of the bands detected by Southern blot analysis and has the following mapping score:
21222212112111222121212111112112112222222
This gave us a map position of 40.90 on chromosome 4S according to the BNL map. At the same position, uaz149(zp19), a 19-kDa zein gene has previously been mapped. Because this position contains most members of the z1A gene subfamily, it corresponds to the z1A-1 location as previously defined (Song, R. and Messing, J, Plant Physiology 130:1626-1635, 2002).
The other group contains a single band (Fig 1, arrow indicated band) and has the following mapping score:
11212221112111212111211121222222122211221
This gave us a map position of 70.20 on chromosome 4S according to the BNL map. At the same position, zpl3a, a zein protein has previously been mapped. This position only contains a few zein gene copies and corresponds to z1A-2 location (Song, R. and Messing, J, Plant Physiology 130:1626-1635, 2002).
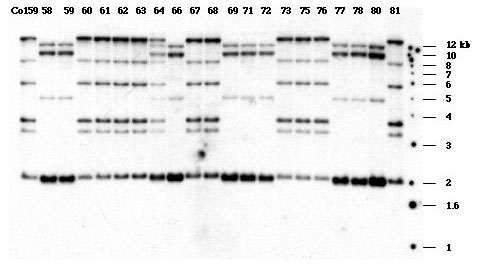
The same approach was taken for mapping the z1B subfamily. In this case, the Tx303xC0159 population cut with HindIII was selected for mapping. To our surprise, only one co-segregating group was detected by Southern blot analysis (Fig 2), with the following score:
11222231221112221112121122221211121112222
This resulted in a map position of 52.30 on chromosome 7S. The same map position is also occupied by uaz68a(zp19) and zpl2b, two alpha zein genes. Even though we found two physically unlinked locations for the z1B subfamily in inbred line B73, they could still be tightly linked in terms of genetic distance because of the low resolution provided by the mapping population (only 41 samples).
A summary of all seven map positions is included in Table 1.
Figure 1. Co-segregation analysis of gene clusters of the z1A subfamily.
Figure 2. Co-segregation analysis of z1B subfamily.
Table 1. Genetic locations of the alpha zein gene family.
| alpha zein subfamily | map position | mapping population | mapping method |
| z1A-1 | chr 4, 40.90 | C0159xTx303 RI | RFLP |
| z1A-2 | chr 4, 72.20 | C0159xTx303 RI | RFLP |
| z1B-1/z1B-2 | chr 7, 52.30 | C0159xTx303 RI | RFLP |
| z1C-1 | chr 4, php200725* | (Mo17xBSSS53)xMo17 | RFLP |
| z1C-2 | chr 4, floury 2* | (Mo17xBSSS53)xMo17 | SNP |
| z1D | chr 1, 122.4; 123.3** | N/A | RFLP |
Return to the MNL 77 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}