Present work in our laboratory aims at extending the methodology of transformation mediated by Agrobacterium tumefaciens to maize genotypes of agronomical value. This includes the identification of reliable protocols aimed to enhance the in vitro responsiveness � related to the development of embryogenic callus cultures � and the frequency of transformation.
In a previous work (Lupotto et al., Maydica 44:211-218, 1999), some parameters for the establishment of a transformation system in maize have been explored in the maize A188 inbred line and in a series of Lo inbred lines of agronomical importance (Bertolini et al., Maydica 36:87-106, 1991). In these latter inbreds, the lack of in vitro responsiveness appeared to be the major bottleneck in obtaining transgenic events. As a first step toward our goal, a series of Lo inbred lines belonging to different heterotic groups (Hartings et al., Maize Coop. Newslet. 76:5-6, 2002) was chosen. Their in vitro response to the embryogenic callus induction (EC) and subsequent regenerative capability were evaluated in the Lo per se and in crosses between each Lo and the highly embryogenic maize model genotype A188 (Table 1). With the exception of Lo1010, the induction of E callus did not appear to be a limiting step; E calli were obtained in all cases, although at different rates depending on the genotype. As expected, the in vitro response of F2 immature embryos derived from each cross showed a significantly higher frequency of EC induction. The most important result was the case of Lo1010xA188, in which a complete switch from non-regenerative to regenerative callus culture was obtained. The number of completely regenerated plants varied largely according to the genotype; in addition, plants raised from each cross were not only higher in number, but also more vigorous, with a properly developed root system established in vitro � which is the most important limiting factor for transplantation into soil � showing a stronger phenotype. Evaluation of the experiment based on the number of completely regenerated plants allowed us to conduct transformations in at least four different crosses.
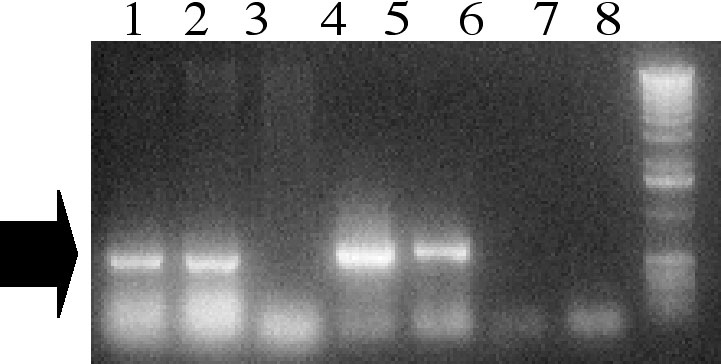
For Agrobacterium-mediated transformation the strain A.tumefaciens EHA105 was used. This was engineered with a binary pCAMBIA3301-based vector, containing the bar gene as selectable marker, and the gus gene as visual marker. The vector also contains the cassette ubi1-b32-nos3� for obtaining the constitutive expression of the maize gene b32 (see above Lanzanova et al.) as a potential anti-fungal gene. Transformation was performed either with immature embryos just after explant or with embryos subjected to a pre-culture period of 5 days. This additional step was introduced with the aim of overcoming the limitation in efficiency due to the deleterious effects of L-cysteine and of the infection process on the subsequent EC induction. As summarised in Table 2, the results obtained strongly supported the adoption of the additional pre-culture. The effect of L-cysteine added during the co-culture period in the case of infection of immature embryos without preculture, varied with the genotype. Of the four crosses evaluated, two were completely inhibited by L-cysteine, while the others were only partially inhibited. In all cases, the absence of L-cysteine raised the rate of EC induction after infection. When a five day pre-culture period before Agrobacterium infection was adopted, the negative effect of L-cysteine on EC induction was drastically reduced; it was also evident that the rates of EC induction paralleled those without L-cysteine. Callus culture in general showed a better phenotype and higher rate of callus proliferation (Figure 1). Presence of the engineered cassette was detected in most of the regenerated plants, as shown by PCR analysis in Figure 2. This result is indicative of the efficacy of the protocol adopted, paving the way for a more consistent set of experiments on the selected maize genotypes. The work is developed within the framework of the EU-funded project SAFEMAIZE (ICA4-CT2000-30033) in FP5.
Table 1.
| Genotype | No. explanted embryos | No. embryogenic calli obtained | % EC | No. regenerating calli | % R | No. plants obtained |
| Lo1096 x A188 | 334 | 266 | 79.6 | 89 | 33.4 | 76 |
| Lo1095 x A188 | 253 | 211 | 83.3 | 118 | 55.9 | 38 |
| Lo904 x A188 | 165 | 124 | 75.1 | 60 | 48.3 | 23 |
| Lo1124 x A188 | 163 | 120 | 73.7 | 17 | 14.1 | 14 |
| Lo1067 x A188 | 192 | 128 | 66.7 | 5 | 3.9 | 2 |
| Lo1066o2x A188 | 196 | 108 | 55.1 | 31 | 28.7 | 29 |
| Lo1010 x A188 | 180 | 131 | 72.7 | 11 | 8.3 | 9 |
| Lo1010 | 200 | 23 | 11.5 | 3 | 13 | 0 |
| Lo1095 | 204 | 134 | 65.7 | 96 | 71.6 | 6 |
| Lo1096 | 192 | 152 | 79.1 | 0 | 0 | 0 |
| Lo904 | 194 | 77 | 39.7 | 12 | 15.6 | 2 |
| Lo1124 | 175 | 97 | 55.4 | 0 | 0 | 0 |
Table 2.
| Genotype | No. explanted embryos | % EC obtained
+cys no cys |
| Infection: T0 | ||
| Lo1096 x A188 | 100 | 0 85.5 |
| Lo1095 x A188 | 100 | 52.8 86.4 |
| Lo904 x A188 | 100 | 40 82.6 |
| Lo1124 x A188 | 100 | 0 53.7 |
| Infection: T5 | ||
| Lo1096 x A188 | 100 | 76.8 84.8 |
| Lo1095 x A188 | 100 | 87.5 67.9 |
| Lo904 x A188 | 100 | 69.5 84.4 |
| Lo1124 x A188 | 100 | 98.8 71.7 |
Figure 1. Embyogenic maize callus in regeneration medium, derived from F2 immature embryos of the cross Lo904 x A188, after Agrobacterium transformation
Figure
2. PCR analysis of T0 events of transformation with Agrobacterium
EHA105(pCAMBIA3301/ubi1-b32). Lanes: 1 to 6 are independent T0 events:
plants 1,2,4 and 5 show the expected band for ubi1-b32; 7. A188 negative
control, 8.Ladder
Return to the MNL 77 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}