The maize endosperm albumin b32 is a Ribosome Inactivating Protein (RIP) normally expressed in the kernel. It has been the subject of extensive studies aimed at investigating and exploiting its action as a defence protein against fungi and insects. Like other RIPs present in the cereal seeds, b32 may play a double role of a storage and a defence protein during seed germination. Ectopic expression of b32 in maize might result in a wider defence action for other tissues. Current work in our laboratories aims to obtain maize plants expressing b32 in various plant parts and during the complete plant growth cycle, with the final goal of verifying its potentiality as a defence gene against fungal pathogens.
A first set of experiments produced a series of transformation events carrying the b32 gene under the 35S CaMV constitutive promoter. Although not specifically suited for monocotiledonous plants, the 35S promoter correctly drives the expression of b32 gene in transgenic wheat and rice (Lupotto, unpubl.). To this purpose, an expression vector carrying the cassette 35S-b32 and ubi1-bar as a selectable marker, was constructed. T0 events, obtained from biolistic transformation of A188XB73 F2 immature embryos, were pollinated with B73 plants to obtain T1 plants, which were grown in a controlled environment (notification B/IT/02/02), during 2002 at the Section of Bergamo. A total number of 344 seedlings were sprayed with L-glufosinate 4g/l at the III-leaf stage. After five days, a visual check of surviving seedlings showed 167 resistant plants and 177 dead plants, thus confirming the expected ratio of 1:1 segregation for Basta resistance. Before Basta treatment, a series of 40 randomly chosen plants were analysed in PCR in order to confirm correspondence between Basta resistance and bar gene presence, and to evaluate the co-presence of both bar and b32 genes. Results obtained confirmed the co-presence of both genes in each resistant plant: 18 plants were Basta resistant and PCR positive, whilst 22 plants died and were PCR negative.
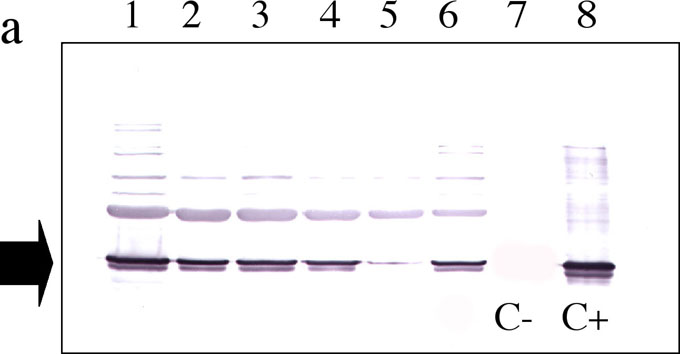
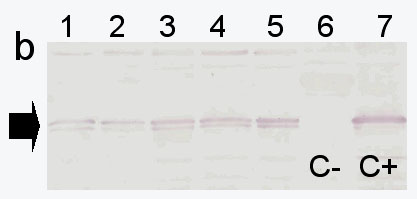
Young plants at the VI-leaf stage were subsequently analysed for control of ectopic b32 expression in the leaf tissue. All b32 PCR+ plants analysed with immunoblots using anti-b32 antibodies - raised in rabbits against purified GST-b32 obtained in E.coli � were positive in western, with the exception of two plants, thus confirming the correct expression of the endosperm protein in leaf tissues (Figure 1a). Plants were grown to complete maturity, were fully fertile and set seeds. Expression of b32 was analysed in adult plants at the stage of flowering, with focus on those tissues more likely to be target sites for fungal invasion: silks, rachis, brace roots and husks. Results obtained gave evidence of expression of the engineered protein in all parts of the b32-engineered adult plants; the two plants which did not express in young leaves were confirmed to be non expressing at later stages (Figure 1b). Current work is focused on b32 expression analysis in germinating seeds and on the interaction between the b32-engineered maize and Fusarium.
The work is developed within the framework of the EU-funded project SAFEMAIZE (ICA4-CT2000-30033) in FP5.
Figure 1. a) Western analysis of T1 plants engineered with 35S-b32, at the VI-leaf stage. Expression was confirmed in all PCR+ plants, except two. The maize control non-transgenic genotype W64A, does not express b32 in leaves (C-) but only in the endosperm (C+). Lanes: 1.SM22, 2. SM30, 3.SM35, 4.SM38, 5.SM2, 6.SM43, 7.W64A leaf, 8. W64A endosperm.
b)
Western analysis for b32 expression in various tissues of plant SM1 at
the flowering stage. Lanes: 1.SM1 silks, 2. SM1 rachis, 3.SM1 brace
roots, 4.SM1 husks, 5.SM1 leaf, 6.W64A leaf, 7. W64A endosperm
Return to the MNL 77 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}