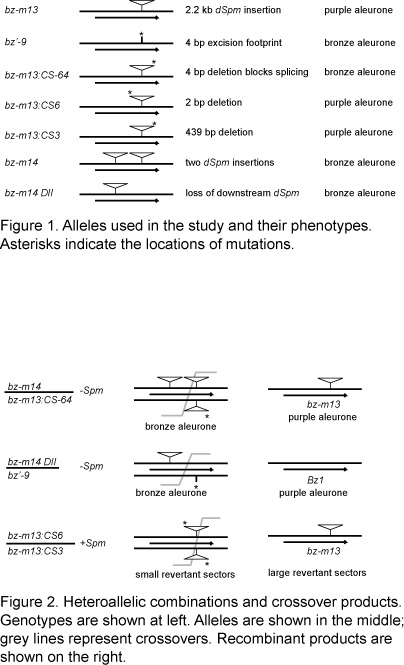
Alleles used in this work were derived from bz1-m13 (Fig. 1). The bz1-m13 allele contains a 2.2 kb defective-Spm (dSpm) element inserted 38 bp downstream of the bz1 intron. In the absence of an autonomous Spm element, seed carrying the bz1-m13 allele have a purple aleurone. The dSpm element is spliced from the transcript using a cryptic 3' splice acceptor site in the dSpm. A partially functional enzyme is made which gives the purple aleurone (Kim, HY et al., PNAS 84:5863, 1987). In the presence of an autonomous Spm element, bz1-m13 is very mutable.
Derivative bz1-m13 alleles obtained in the presence of an autonomous Spm element are easily identified by their phenotype, and a number of derivatives have been isolated and characterized. bz1-m13:CS6 (Schiefelbein, JW et al., in Plant Transposable Elements pp 261, 1988) and bz1-m13:CS3 (Okagaki, RJ et al., Plant Cell 4:1453, 1992) are deletion derivatives of bz1-m13. bz1-m13:CS64 (Okagaki, RJ et al., Plant Cell 4:1453, 1992) has a four basepair deletion that disrupts the cryptic 3' splice acceptor site; this allele gives a bronze colored aleurone. In bz1'-9, the dSpm insertion excised leaving behind a four basepair footprint (Okagaki, RJ et al., MGNL 64:83, 1990).
bz1-m14 was derived from bz1-m13 and has two dSpm insertions. Its structure is consistent with a replicative transposition of the dSpm element in bz1-m13 back into the bz1 locus (Sullivan, TD unpublished data). The upstream element is unique to bz1-m14, and the downstream element is in common with bz1-m13. bz1-m14DII has lost the downstream dSpm element; excision either restored the non-mutant sequences or the transposable element footprint restored the correct reading frame (Sullivan, TD unpublished data).
The first experiment looked for recombination in two heteroallelic combinations where the mutations were separated either by approximately 500 bp or by 2.7 kb; the difference was due to the presence of the dSpm insertion (Fig. 2). A recombination event between the dSpm insertion in bz1-m14DII and the excision footprint in bz1'-9 would produce a functional Bz1 allele and give a purple aleurone. Similarly, a recombination event occuring between the upstream dSpm insertion in bz1-m14 and the deletion in bz1-m13:CS64 would produce a functional bz1-m13 allele and give a purple aleurone. In the bz1-m14DII/bz1'-9 combination this recombination event would occur in a region approximately 500 bp in length while the interval in the bz1-m14/bz1-m13:CS64 combination was approximately 2.7 kb long. If the repetitive dSpm sequence does not participate in recombination the reversion frequency should be similar. But if the dSpm sequence participates in recombination revertants should be five times more frequent from the bz1-m14/bz1-m13:CS64 combination assuming that reversion frequency is proportional to the physical distance between mutations.
Problems with nicking prevented us from making crosses with the bz1 tester. Plants were selfed instead. Twelve purple revertant kernels were found in 65,700 selfed seed from bz1-m14DII/bz1'-9 plants. Six purple revertant seeds were found in 58,500 selfed seeds from bz1-m14/bz1-m13:CS64 plants. This result suggests that a dSpm element within a gene does not participate in recombination and may even inhibit recombination in adjacent sequences. The two dSpm sequences in bz1-m14 may pair; this could inhibit the recovery of revertants. We hope to isolate new derivative alleles from bz1-m14 and bz1-m14DII that would lack the upstream insertion.
The second experiment looked for recombination within the dSpm sequence to generate an intact bz1-m13 allele that could be identified by its mutable phenotype. The deletion derivative bz1-m13:CS6 has a 2 bp deletion at the 5' end of the element as it is oriented in bz1. bz1-m13:CS3 has a 439 bp deletion at the 3' end of the element. A crossover within the 1.8 kb of overlapping dSpm sequence would produce the bz1-m13 structure. In the presence of the autonomous element Spm, bz1-m13:CS3 and bz1-m13:CS6 give small revertant sectors. bz1-m13 +Spm gives early frequent revertant sectors. bz1-m13:CS3/bz1-m13:CS6 plants were crossed with a sh bz1 tester homozygous for the autonomous Spm element, and ears were examined for kernels with large revertant sectors. Weather and worms hit this field hard, and this experiment will be repeated.
Return to the MNL 76 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}