In the hybrids between tetraploid maize and T.dactyloides whose offspring we are continuing to investigate, Laikova reported that 56-chromosome F1 hybrids (20Zm + 36Td) were genetically heterogeneous (In: Petrov, DF (ed.)Apomixis and its role in evolution and breeding, New Dehli, India, 79-87, 1984). Hybrids H416 and H418 (2n=56), developed from backcrossing apomictic F1 hybrid H278 (2n=56) with maize, were also the apomicts and had embryo sacs similar to those observed in the paternal Tripsacum. At the same time among F1 hybrid H166 (2n=56) only BII-hybrids with 38 chromosomes (20Zm + 18Td) were recovered in the BC1. Sexual reproduction was confirmed by the presence of meiotically derived linear tetrads of megaspores.
Offspring of the maize-Tripsacum hybrids H278 and H32 (2n=56; 20Zm + 36Td), 39-chromosome forms (30Zm + 9Td) retained apomictic reproduction (Sokolov, VA et al., Russian Genetics 34:499-506, 1998). Low female fertility in the F1 hybrids was overcome in subsequent generations by reducing the number of Tripsacum chromosomes and selecting the most productive genotypes. However, low seed production remained a problem. The practical use of the hybrids was also restrained by their male sterility. It was important to understand the mechanism of apomixis inheritance when genes of eastern gamagrass were expressed in cytoplasm of maize in the presence of three maize genomes (thirty chromosomes) and only nine Tripsacum chromosomes. To determine the causes of decreased kernel set, we examined megasporogenesis, megagametogenesis and pollen tube growth in the maize-Tripsacum hybrids. This was in collaboration with scientists from the Komarov Botanical Institute, Saint Petersburg, Russia, and Plant Research International, Wageningen, the Netherlands.
For the study of megasporogenesis and megagametogenesis, ears 3-10 cm long were fixed in FAA [3:7:1 (V:V:V) formalin: 70% alcohol: glacial acetic acid] for 48 hours. The fixed material was stored in 70%-ethanol. The ovules were cleared in methyl benzoate and examined using phase-contrast microscopy and Nomarsky optics (Naumova, TN et al., Sex. Plant Reprod. 12:43-52, 1999).
To study the pollen tube growth, ears were pollinated by maize pollen and then fixed in FAA two days after pollination (DAP). The styles were placed in 10% KOH for 1.5 hours. After maceration the styles were washed with distilled water, placed on slides and stained in a drop of 0.005% aniline blue in 0.15M K2HPO4. The styles were squashed and observed for the presence of the pollen tubes using fluorescent microscopy.
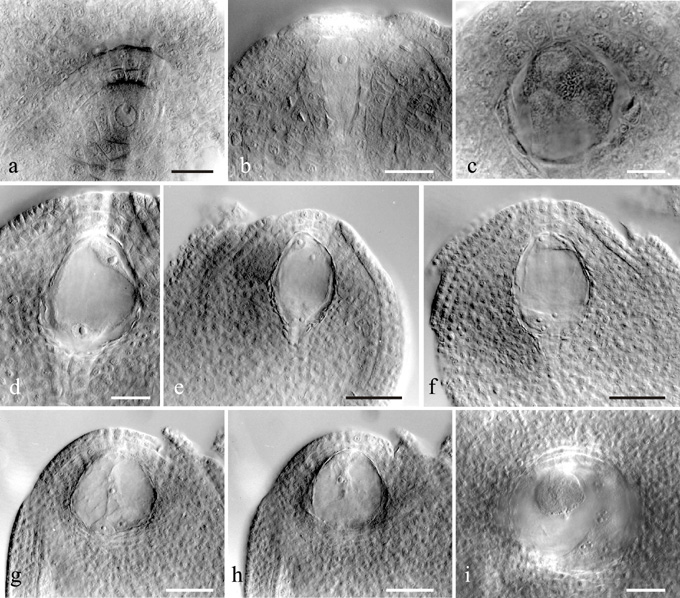
More than 120 ovules from 12 plants of two different lines 4x-6 and W39 with 30 chromosomes of maize and 9 chromosomes of Tripsacum were examined. Megasporogenesis and embryo sac formation in both lines proceeded similarly. The subepidermal nucellar cell enlarged and became the archesporial cell and later differentiated into the megaspore mother cell (MMC) (Fig. 1a). Instead of initiating meiosis, characteristic of sexual plants, the elongated MMC enlarged and became vacuolated, the nucleus remained inactive, the chalazal end of the cell became narrow. The thickening of the cell wall was accompanied by degeneration of the adjacent nucellar cells. Integuments enclosed the micropylar part of the ovule (Fig. 1b). After isolation, MMC divided mitotically (Fig. 1c) to produce a 2-nucleate megagametophyte with a huge central vacuole (Fig.1d). The second mitotic division produced a 4-nucleate embryo sac (Fig. 1e) and the third division produced an 8-nucleate embryo sac (Fig. 1f). After cellularization, the embryo sac consisted of two synergids, egg-cell, two polar nuclei and three antipodal cells, which often divided later on to form five or six antipodals (Fig.1g, h). The complete absence of meiosis and the megaspore tetrad formation indicated that the 39-chromosome maize-Tripsacum hybrids reproduced by mitotic diplospory and the Antennaria-type megagametogenesis. In the material studied, neither meiosis nor abnormal embryo sac development were observed.
The embryo in the maize-Tripsacum hybrids developed parthenogenetically. Fertilization of polar nuclei is necessary for the formation of the endosperm. In the isolated ears that were not pollinated, only unfertilized embryo sacs with enlarged egg-cell, fused polar nuclei and multiplying antipodals were found on the second to third day after the ejection of stigmata. Thus, pseudogamous apomixis occurred in the 39-chromosome maize-Tripsacum hybrids. This is similar to the development of eastern gamagrass as reported by Farquharson, LI (Am.J.Bot.42:737-743,1955), Brown, WV and Emery, WHP (Am.J.Bot. 45:253-263,1958) and Burson, BL et al.(1990).
Cases of embryo development without pollination were not observed in our maize-Tripsacum hybrids. Moreover, at 3DAP the endosperm usually overtook the egg cell in the rate of cell divisions (Fig. 1i). Pollination and fertilization of the polar nuclei appeared to be the signal of parthenogenetic embryo development. A similar phenomenon was described in apomictic Arabis holboellii (Naumova, TN et al., Sex. Plant Reprod. 2001, in press). Parthenogenesis without pollination was reported by Laikova, LI (1984) and Fokina, ES (In: Petrov, DF (ed.) Inducirovannyi mutagenez i apomiksis, Novosibirsk, USSR, pp. 57-63, 1980).
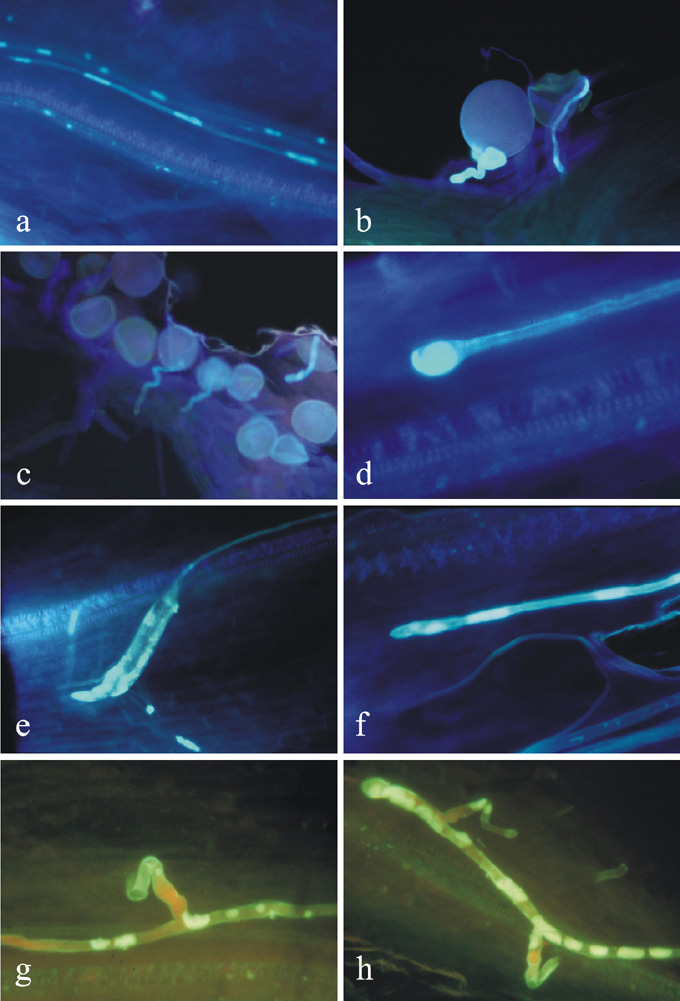
The absence of embryo and endosperm development in part of the ovules in the pollinated ears suggested that fertilization of polar nuclei did not occur in these. To elucidate the causes of this phenomenon, we examined the pollen tube growth in line 4x-6 (30Zm + 9Td chromosomes) in 1999 and conducted a comparative study of this line with the 38-chromosome line (20Zm + 18Td) in 2000. The investigation of styles of pollinated ears showed that in rare cases, the maize pollen tubes had regular callose plugs and grew toward the ovule (Fig. 2a). More often the pollen tubes stopped growing shortly after germination (Fig. 2b, c) and sometimes within the stylar tissue (Fig. 2d, e, f). Lack and stoppage of growth were accompanied by anomalous callose deposition in the tubes, tube twisting and branching (Fig. 2g, h). In 1999, after the 39-chromosome hybrids were pollinated with the maize Tetraploid Shumnogo (2n=4x=40), pollen tubes grew toward the ovule in 15% of the styles, whereas in the maize control pollen tube growth was 100%. The number of pollen tubes reaching the ovule was also considerably lower in the hybrids when compared to maize. There were 1-2 tubes per style in hybrids as compared to 7-9 tubes in maize. In 2000, we also studied the influence of the pollinator on the pollen tube growth in the 39- and 38-chromosome hybrids. Diploid (Popcorn) and tetraploid (Tetraploid Shumnogo, Purple marker) maize were used as the pollen sources. For each combination, 10 ears were pollinated and 40-50 styles from each ear were analyzed. The percentage of styles with pollen tubes that reached the ovule was calculated. Significant differences for this characteristic between pollinators and the maternal genotypes were not observed. The mean values of pollen tube growth were higher than in 1999: Popcorn - 41%, Tetraploid Shumnogo - 48%, Purple marker - 32% (P>0.05). However when analyzing the material by pollination day it was established that in the last three days of pollination, beginning from 27 August, the percent of styles with tubes and the number of tubes per style increased (Table 1).
Table 1. Pollen tube growth in the maize-Tripsacum hybrids on
different days of pollination.
| Pollination date | 10.08 | 15.08 | 24.08 | 25.08 | 26.08 | 27.08 | 28.08 | 29.08 |
| Number of styles studied | 78 | 88 | 116 | 412 | 328 | 550 | 121 | 190 |
| Number of styles with pollen tubes | 32 | 12 | 19 | 109 | 53 | 374 | 80 | 149 |
| % of styles with tubes | 41 | 4 | 16 | 27 | 17 | ***68 | ***66 | ***78 |
| Number of tubes found | 62 | 18 | 34 | 201 | 92 | 769 | 193 | 499 |
| Number of tubes per 1 style | 0.80 | 0.20 | 0.29 | 0.49 | 0.28 | ***1.40 | ***1.60 | ***2.63 |
Note: *** - the differences between days 27-29.08. and the other days of pollination are significant with P<0.001
The weather conditions in the pollination period were characterized by a lack of sharp fluctuations with 15-20C in the day time and 8-9C at night on average. However, on the night of 26-27 August a fall of temperature to 1-2C occurred, so differences between the average and decreased temperature was about 7 degrees. As known from the literature, a change in temperature adversely affects the pollen fertility, the process of fertilization and ovary development (Jakobsen, HB and Martens, H, Annals Bot. 74(5):493-501, 1994; McKee, J and Richards, AJ, Annals Bot. 82(3):359-374, 1998; Tromp, J and Borsboom, O, Scientia Horticulturae 60(1-2):23-30, 1994). It is possible that the low temperature influenced incompatibility mechanisms in the maize-Tripsacum hybrids. Apparently in this case the short-term decrease in temperature weakened the incompatibility of maize pollen tubes and tissues of style of the maize-Tripsacum hybrids without having an effect on viability of pollinator-maize pollen. The effect of temperature shock on incompatibility has been reported in Lolium perenne (Wilkins, PW and Thorogood, D, Euphytica 64:65-69, 1992).
Thus, one can conclude that abnormal growth of maize pollen tubes in styles of 39- and 38-chromosome maize-Tripsacum hybrids was one of the main reasons for poor kernel set. The three maize-pollinator varieties used did not significantly influence the efficiency of pollination. A short-term low-temperature shock promoted the weakening of the incompatibility mechanisms.
The authors thank B.L Burson for the fruitful discussions and K. Boutilier for comments on the manuscript. The research was supported by NWO, the Netherlands, Research Grant No.047.007.019 and Russian Foundation of Basic Research, Grant No.00-04-49542.
Figure 1 (a) Ovule with archesporial cell. (b) Ovule with big megasporocyte (MMC) showing vacuolization and tendency to dyplospory. (c) Ovule with diplosporous one-nucleate embryo sac of Antennaria-type originated directly from MMC. (d) Two-nucleate embryo sac. (e) Four-nucleate embryo sac. (f) Eight-nucleate embryo sac prior to the cell formation. (g, h) Eight-nucleate embryo sac after cellularization and with two synergids, egg cell, central cell with two unfused polar nuclei and three antipodal cells (two successive focus levels of the same ovule). (i) Egg cell at the metaphase stage and 4-nuclei endosperm. Bar: for a, i, 50 µm; c, 20 µm; b, d, e, f, g, h, 100 µm.
Figure
2. (a) Pollen tube growth is normal, the plugs are equi-distant from
each other. (b, c) Pollen tube growth is interrupted at an early stage,
soon after germination. (d, e, f) Pollen tube growth is interrupted at
later developmental stage, inside the style. (g, h) Abnormality (branching)
of the pollen tubes inside the style.
Return to the MNL 76 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}