*also affiliated with Dpto. de Ciencias Biológicas, FCEN, UBA.
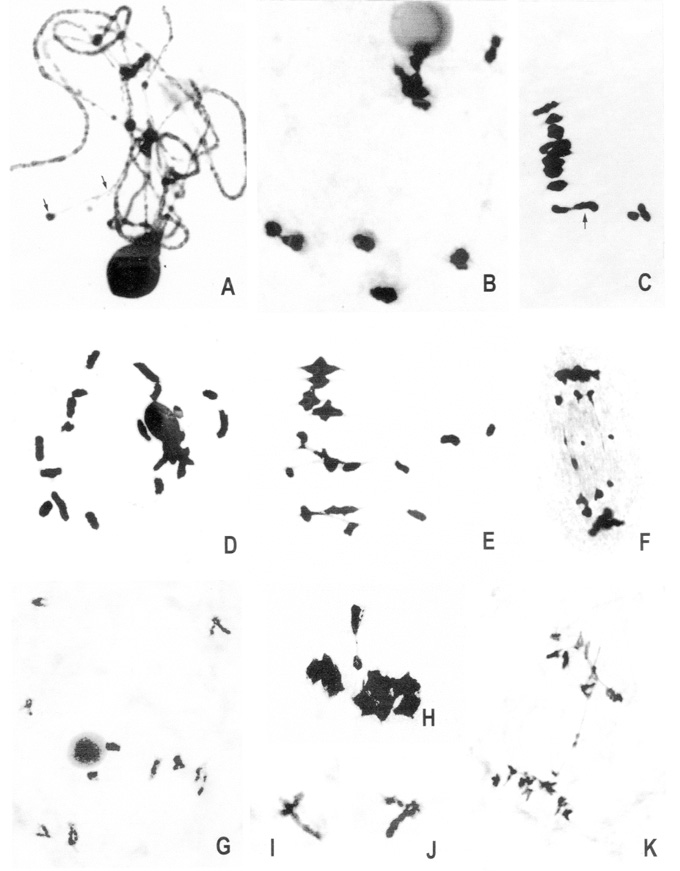
The F1 hybrid Z. luxurians x Z. mays ssp. parviglumis showed pachytene heterozygosis in the knob regions (Fig. 1A), meiotic asynchrony of two groups of 5 II each (diplotene-diakinesis) (Fig. 1B), heteromorphic bivalents and univalents of different size (diplotene-diakinesis, metaphase I) (Fig. 1C), bridges and fragments (anaphase I), and laggard chromosomes (anaphase I and II). These phenomena were also observed in the F1 hybrid Z. luxurians x Z. mays ssp. mexicana (Fig. 1D, E, F)). In the F1 hybrid Z. luxurians x Z. mays ssp. mays heteromorphic bivalents and 1-2 bridges and fragments (anaphase I) were observed (Fig. 1 G-K).
In these hybrids 8 II + 4 I was the most frequent configuration, although the range of univalents were 0-20 per cell (Fig. 1D). Moreover, they were highly sterile (pollen stainability: 0-12%) and showed loss of pairing in heterozygotes for knobs in pachytene. The genome size varies between 2C= 5.8 - 6.8 pg in the subspecies of Zea mays, while Z. luxurians has 2C= 9 pg. The difference in the genome size between the hybrid�s parental species could explain the presence of heteromorphic bivalents. The presence of two asynchronic groups of 5 II was also frequent.
The bridges and fragments, observed in anaphase I, in the studied hybrids would indicate that the parental species differ in 2-3 paracentric inversions between them.
The presence of two groups of 5 II suggests that the genomes of these hybrids maintain structural and functional independent domains, despite the existence of intergenomic rearrangements revealed by molecular data.
Figure 1. A-C: Z. luxurians x Z. mays ssp. parviglumis. D-F: Z. luxurians x Z. mays ssp. mexicana. G-K: Z. luxurians x Z. mays ssp. mays.
A: Pachytene: the arrows show irregular pairing in the knob regions.
B: Diakinesis: meiotic asynchrony of two groups of 5 II each.
C. Metaphase I: the arrow shows a heteromorphic bivalent, univalents
of different size are observed off the plate.
D: Diakinesis: 20 univalents.
E: Metaphase I: heteromorphic bivalent and univalents of different
size off the plate.
F: Anaphase I: Two bridges and two fragments.
G: Diakinesis: 10 heteromorphic bivalents.
H: Metaphase I: heteromorphic bivalent.
I - J: Open heteromorphic bivalents.
K: Anaphase I showing one bridge.
Return to the MNL 76 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}