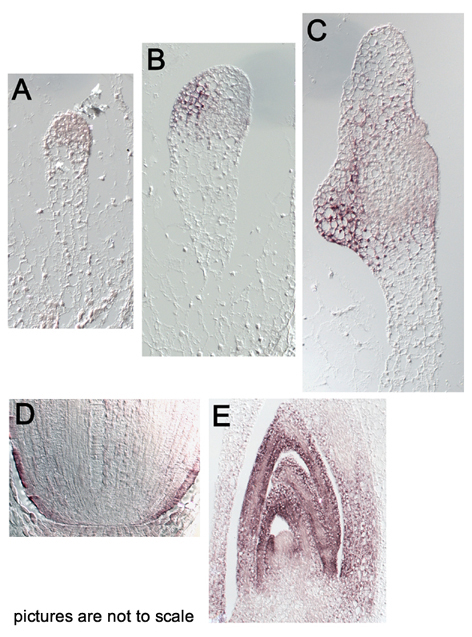
Sh1 transcripts are absent, or below the detection level in the embryo proper during the proembryo-stage (Fig. A). In the transition-stage embryo, Sh1 transcripts accumulate in the abaxial hemisphere of the embryo-proper. These Sh1 expressing cells contribute to the development of the scutellum and rapidly enlarge at the end of the transition stage. This cellular enlargement is the main mechanism responsible for the outgrowth of the scutellum (Elster et al., Dev. Genes. Evol. 210:300-310, 2000). In contrast, cells located adaxially in the region of the SAM-anlage stay compact and remain free of Sh1 activity (Fig. B). A series of longitudinal sections through coleoptilar-stage embryos show that the Sh1 transcriptional activity is not equally distributed throughout the scutellum, but a patchy pattern appears to be associated with domains of rapid cellular enlargement (Fig. C).
At later embryonic stages, Sh1 is downregulated in the elaborated scutellum, whereas expression at high levels is now detected in the root/shoot axis. Strong expression is found in the rhizodermis, but lower Sh1 transcript levels are observed in the central cylinder of the root. No Sh1 transcripts are observed in the root meristem (Fig. D), which is similar to the embryonic shoot apical meristem. Both primary meristems are therefore essentially free of Sh1 activity although cells in the meristems strictly depend on the import of carbohydrates.
In the shoot, high Sh1 transcript levels are found in the epidermal layer of developing leaves. This Sh1 activity in embryonic leaves is not restricted to the epidermis but extends into the parenchymal cell layer, although at a lower level relative to that in the epidermal layer, additionally weak expression can be detected in expanding coleoptile (Fig. E). Activation of the Sh1 promoter in maize is thus linked to the initiation of leaf primordia as has been described for the tomato SAM (Pien et al., Plant J. 25:663-674, 2001).
In conclusion, transcriptional activity of the Sh1 gene is highly
dynamic during embryogenesis. The absence of Sh1 transcripts in
the primary root and shoot meristems with high rates of cell proliferation
argues against a simple sink-based control. In contrast, the obvious link
between cellular enlargement and Sh1 transcription during scutellum
development, supports a direct contribution to cell wall synthesis (Amor
et al., Proc. Natl. Acad. Sci. USA 92:9353-9357, 1995), an assumption compatible
with the repression of Sh1 promoter activity by the cellulose biosynthesis
inhibitor dichlorobenzonitril (Maas et al., EMBO J. 11:3447-345, 1990).
One interesting new aspect of the Sh1 pattern is the preferentially
high activity in the epidermal cells of the primary root and young leaves,
which may be related to functional specificities of both outer cell layers.
Return to the MNL 76 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}