Unlike wheat, many maize inbred lines show significant levels of resistance to WSMV. There are three known genes in maize for resistance to WSMV located on three different chromosomes (McMullen, MD et al., Mol. Plant-Microbe Interact. 7:708-712, 1994). The wsm1 gene was found to map to chromosome 6S between the maize restriction fragment length polymorphism (RFLP) loci umc85 and npi235, near the nuclear organizer region (NOR). Another viral resistance gene, mdm1, conferring resistance to maize dwarf mosaic virus (MDMV) was closely linked on chromosome 6. The long arm of maize chromosome 10 harbored the wsm3 locus. Mapping experiments placed wsm3 between markers umc163 and umc44. Wsm2 was positioned on the maize genetic map near the centromere of chromosome 3 between umc102 and umc18. This map position is near the rp3 locus, a dominant gene for resistance to Puccinia sorghi, the causal agent of maize common rust. Genes conferring resistance to maize mosaic virus (MMV) and sugarcane mosaic virus (SCMV) and QTLs for resistance to other pests also map to this general area (McMullen and Simcox, Mol. Plant-Microbe Interact. 8:811-815, 1995; Ming et al., Theor. Appl. Genet. 95:271-275, 1997; Xu et al., Mol. Gen. Genet. 261:574-581,1999)
A recent attempt to identify candidate sequences for resistance genes identified the PIC13 probe which hybridizes to a family of resistance gene-like sequences at the Rp3 locus (Collins et al. Mol. Plant-Microbe Interact. 11:968-978,1998). Recently we examined the relative position of the rp3 and wsm2 loci to determine if the PIC13 gene family might also include the Wsm2 gene. A line carrying the Rp3-A allele in an H95 genetic background (Rp3-A/Rp3-A, Wsm2/Wsm2), resistant to the rust (Puccinia sorghi) biotype IN1 and the Wooster WSMV strain was developed. This line was crossed to the rust and WSMV susceptible inbred OH28 (rp3/rp3, wms2/wsm2). The F1 was backcrossed to the susceptible OH28 parent and BC1 progenies self-fertilized. Individual seedlings of the resulting BC1F1 families were scored in the greenhouse after inoculation with talc-diluted rust spores and rub inoculation with the Wooster WSMV strain (Louie, R, Phytopathology 76:769-773, 1986). Plants with resistance to WSMV are symptomless. However, a �virus resistant� recombinant class would be the most likely class in which escapes could occur and would be the most difficult to score. To reduce the number of escapes, a minimum of three independent virus inoculations were made, each spaced 2-3 days apart, on the two/three leaf-stage seedlings. Furthermore, efforts were made to maintain a high virus titer in our inoculum and to introduce an equal amount of inoculum load to each seedling.
Resistance or susceptibility to rust infection was very definitive and was observed as either hypersensitive flecking or sporulation, respectively. We observed no latent symptoms. One application of fresh rust urediniospores was made to the seedling leaves, typically after the second virus inoculation. The scoring of rust and WSMV symptoms commenced seven days post-inoculation. Recombinants were identified as being either rust resistant/virus susceptible (RRVS) or rust susceptible/virus resistant (RSVR). Table 1 summarizes these scoring data. The recombinant heterozygous seedlings were transplanted from flats to five-gallon pots and then self-fertilized. Self-fertilized families from most of the putative recombinants were progeny tested. Progeny testing indicated that all but three individuals assigned to the RRVS class had been scored correctly. However, individuals in the RSVR class were more often mis-scored. Even after progeny testing, a disproportional number of what appeared to be RSVR-recombinant types resulted. Relatively equal proportions of the two recombinant classes are expected from such an experiment. There are two explanations that could account for these observations. First, there remains the possibility that poorly challenged plants or latent viral symptoms led to mis-classification of virus resistant plants even during progeny testing. Alternatively, it is possible that there is a second, unidentified gene segregating in this population. The presence of a second gene may make a wsm2/wsm2 plant appear to be resistant to WSMV, and lead to the inflated numbers we observed in the virus resistant class.
Because we were much more confident in accurately scoring them with few escapes or mis-scores, we chose to use the frequency of the RRVS-class seedlings to calculate map distance. This gave an estimate of roughly 3 cM between the rp3 and wsm2 loci.
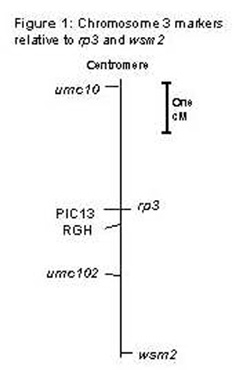
The recombinant individuals were also used to order DNA markers in the rp3-wsm2 interval. DNAs were analyzed from 29 progeny-tested, homozygous individuals and 20 other recombinants that were not homozygous. The PIC13 probe detected a single polymorphic HpaII restriction fragment and two polymorphic EcoR1 restriction fragments. All three polymorphisms cosegregated perfectly with each other and with rp3. This indicates that at least most of the PIC13 family members cluster tightly to the rp3 locus and that it is unlikely that any members of this family account for the Wsm2 gene. A second resistance gene analog clone, designated RGH, was obtained from M. Chen and J. Bennetzen. RGH was placed between PIC13 and wsm2 on this population, more tightly linked to PIC13 than to wsm2. The core markers umc10 and umc102 were also mapped with this population and their positions are in agreement with the UMC 1988 molecular marker map of maize chromosome 3 (Davis, G et al., MNL 72:118-128, 1998). Figure 1 shows the position of molecular markers for this chromosomal region in relation to rp3 and wsm2. The Mv1 gene for resistance to MMV was placed 4 cM below umc102 (Ming et al., Theor. Appl. Genet. 95:271-275, 1997) and may be close to the Wsm2 locus. The Scm2 gene, conferring resistance to SCMV, was mapped approximately 4 cM above umc102 (Xu et al., Mol. Gen. Genet. 261:574-581,1999), and may be close to Rp3. The PIC13 and RGH probes should be mapped with respect to Scm2 to scrutinize these resistance gene-like sequences as candidates for this gene.
Table 1. wsm2 mapping population:
greenhouse data
| Non-recombinant phenotypes | Originally identified |
| RSVS | 403 |
| RRVR | 1,348 |
| Recombinant phenotypes | |
| RRVS | 29 |
| RSVR | 69 |
| Population total | 1,849 |
Return to the MNL 75 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}