Low and high WER content in the lines obtained under such selection (designated as LFWER and HFWER lines with o2-lf; Bg-lf and o2-hf; Bg-hf genotypes, respectively) was determined by change in state in the mutable responsive allele o2-m(r):3449 and the regulatory element Bg-3449. Newly arisen alleles o2-lf and o2-hf were characterized by low and high rate of rbg excision respectively. The regulatory element Bg-hf contained in HFWER lines differed sharply from the Bg-lf in its ability to cause rbg excision at different doses (MNL 73:76-79; Maydica 44:195-203, 1999).
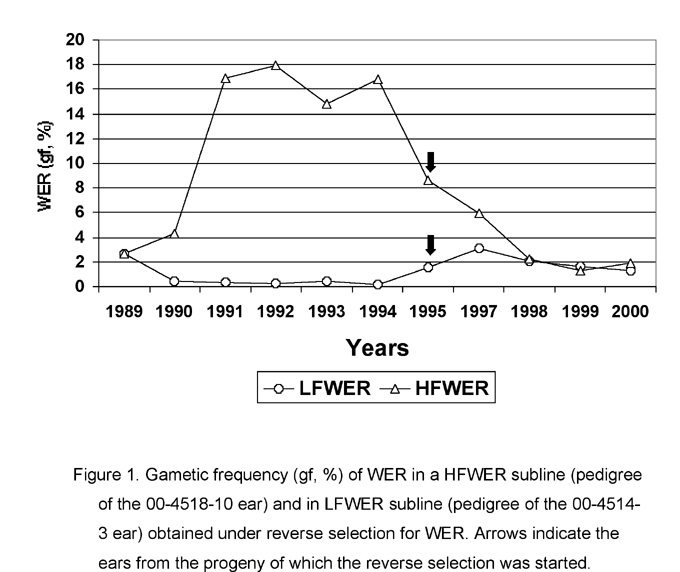
The efficiency of reverse selection for WER content. In 1997 (on the material obtained in 1995) we started the reverse selection for WER content (i. e. selection for WER increase in LFWER and WER decrease in HFWER lines). Results obtained after two years of selection indicated its effectiveness for WER decrease in HFWER lines and ineffectiveness for WER increase in LFWER lines (Maydica 44:195-203). Further selection confirmed these conclusions (Fig. 1). The higher stability of the o2-lf allele in relation to frequency of rbg excision indicates the lower level of organization of this allele in comparison to o2-m(r):3449 and o2-hf alleles. Higher stability of o2-lf could be connected with the loss of some of its genetic properties, resembling in this respect the higher stability of the o2 locus after the reversion of the mutable allele at this locus (Salamini, Cold Spring Harbor Symp. Quant. Bio. 45:467-476, 1981). Since the o2-lf retains its ability for reversion to normal we can assume that this loss did not affect the o2 locus, but was caused by deletion in the rbg element contained in the o2-lf allele. In this connection it is necessary to mention that internal deletions in the nonautonomous (receptor) dSpm element of the Spm/En system can delay frequency and timing of excision of this element (see for example Schiefelbein et al., Proc. Natl. Acad. Sci. USA 82:4783-4787, 1985; Schwarz-Sommer et al., EMBO J. 4:2439-2443, 1985).
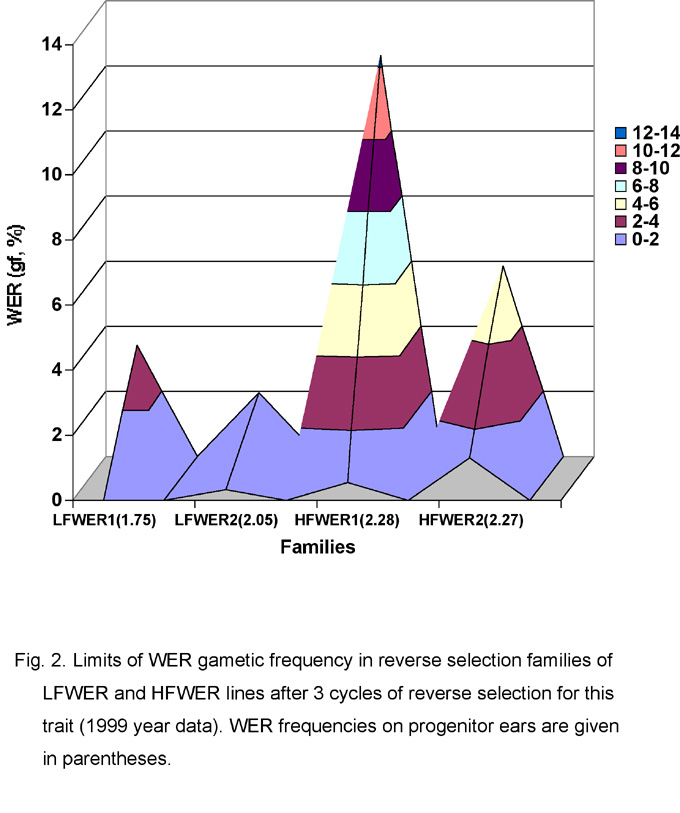
Low frequency of whole endosperm revertants can be conditioned by different genetic mechanisms. During reverse selection for WER frequency, it was observed that the limits of variation of WER frequency in the progeny of the HFWER ears with low revertant content equal to that of the LFWER lines were much higher in comparison with the variation of the same trait in LFWER families. Thus, after 3 cycles of reverse selection, variation of WER frequency in 2 HFWER reverse selection families was in the range of 0.56-12.31% (gametic frequency of WER in the ears from which they were originated was 2.27 and 2.28%). Variation of the same trait in reverse selection progenies of LFWER sublines was equal to 0-3.43% (these families originated from the ears with gametic WER frequency of 1.75 and 2.05%) (Fig. 2). Differences in variability of WER content could reflect different mechanisms of the control of low frequency of the rbg excision from the mutable alleles of the LFWER lines and the HFWER reverse selection sublines.
Expressivity of quantitative traits in lines differing with states of the Bg-rbg system components. We reported that lines obtained under disruptive selection for WER differed also in some quantitative traits: lines obtained under selection for high revertant content had higher kernel weight and volume, number of leaves, longer period from emergence to flowering (MNL 73:76-79; Maydica 44:195-203) and were more sensitive to herbicide application (MNL 74:57-58) in comparison to the lines obtained under selection for low revertant frequency.
We also observed that sensitivity to herbicide "Buctril D" in the lines obtained did not depend from the presence of the active regulatory element in these lines (MNL 74: 57-58).
The mentioned differences of kernel weight and volume were observed on variegated kernels of the forms containing active regulatory elements. On the material obtained in 1999, we analyzed kernel weight and volume in LFWER and HFWER lines and their derivatives that lacked regulatory elements (o2-lf; +Bg and o2-hf; +Bg derivatives, respectively). (The methods of the analysis were described earlier (MNL 73:76-79)). Data obtained showed that superiority in kernel weight and volume of the lines obtained under selection for high revertant frequency was observed both on variegated and non-variegated kernels the latter formed on sublines lacking regulatory elements (Table 1).
Higher values of kernel density observed with the o2-lf; +Bg derivatives most likely resulted from the action of polygenic modifiers which conditioned higher endosperm vitreousity. These modifiers apparently also lessened the differences in kernel weight between the LFWER and HFWER derivatives lacking regulatory elements. The results obtained indicate that the differences of the studied quantitative traits do not depend directly on WER content and on the presence of active regulatory elements in the lines studied.
Table 1. Values of some characteristics
of variegated and non-variegated kernels of the lines with low and high
WER frequency (with o2-lf; Bg-lf and o2-hf; Bg-hf genotypes,
respectively) and their derivatives lacking regulatory elements (o2-lf;
+Bg and o2-hf; +Bg genotypes, respectively).
| Genotypes | Ears analyzed | 50 kernel weight, g | 50 kernel volume, cm3 | Kernel density, g/cm3 |
|
|
||||
| o2-lf; Bg-lf | 7 | 5.83a* | 4.40a | 1.326a |
| o2-hf; Bg-hf | 8 | 7.05b | 5.21b | 1.354a |
| o2-hf; Bg-hf as % of o2-lf; Bg-lf | 120.9 | 118.4 | 102.1 | |
|
|
||||
| o2-lf; +Bg | 14 | 5.89a | 4.69c | 1.257b |
| o2-hf; +Bg | 16 | 6.69b | 5.50b | 1.216c |
| o2-hf; +Bg as % of o2-lf; +Bg | 113.6 | 117.3 | 96.7 | |
* For each trait a common letter at
the means indicates insignificance of the differences between them (P£0.05).
Return to the MNL 75 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}