The aim of the present work was to verify our hypothesis about the possible involvement of the glutathione system in the redox regulation of transcriptional activity in mitochondria.
Mitochondria were prepared from 3-day-old etiolated maize seedlings of hybrid VIR42 MV. The isolation of mitochondria and assay of RNA synthesis in organello are described in our accompanying note.
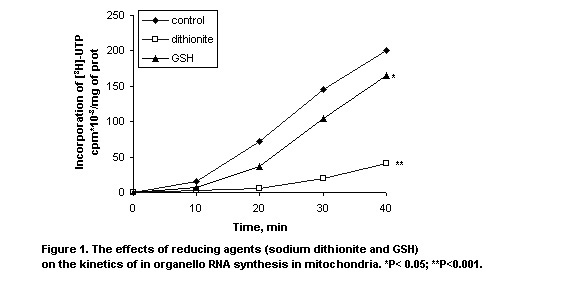
The effects of the reduced forms of glutathione and sodium dithionite on the kinetics of RNA synthesis in maize seedling mitochondria are shown in Figure 1. The mitochondrial transcriptional activity is seen to decrease in the presence of the reduced glutathione. The effects of reduced glutathione and sodium dithionite on the activity of mitochondrial RNA synthesis were similar to those observed for the protein synthesis in mitochondria in the presence of these reduced agents (MNL 72:33, 1998).
We reported previously (MNL 69:63-64, 1995; MNL 70:29-30, 1996) that under oxidizing conditions mitochondrial transcription and translation were activated, while under reducing conditions they were strongly repressed. Experimental study of redox conditions impact on the activity of mitochondrial DNA topoisomerase I in maize showed that under oxidizing conditions in the presence of GSSG a significant decrease of topoisomerase activity was observed, whereas under reducing conditions in the presence of GSH enzyme activation was observed (MNL 73:39-40, 1999). This raises the question of whether there is a functional relation of the changes in the mitochondrial DNA topoisomerase I activity in oxidizing and reducing conditions with the phenomenon of changes in the transcriptional activity of the mitochondrial genome in the same conditions.
It was demonstrated previously with the use of a reconstituted system that human topoisomerase I can serve as a repressor of basal transcription (Chen and Xu, Biochem. Mol. Biol. Intern. 39:941-948, 1996). This repression can be overcome by transcriptional activators or TFIIA. It was also reported that a repressing effect of human topoisomerase I was observed only in TATA-box-containing promoters and was mediated by the TATA-binding protein (Chen and Xu, Biochem. Mol. Biol. Intern. 39:941-948, 1996). By analogy with the case described, we presume that the plant mitochondrial topoisomerase I is able to repress the transcription of all or a part of mitochondrial genes. In this case the repression of its activity by oxidizing conditions result in an enhancement of mitochondrial transcription. The likelihood of such a situation is supported by an earlier observation of enhanced transcription by isolated maize mitochondria under oxidizing conditions (MNL 69:63-64, 1995). Under reducing conditions the mitochondrial DNA topoisomerase I is activated and represses the transcriptional activity of mitochondria. We suggest that DNA topoisomerase I can be a regulator of the expression of all or a part of the genes in mitochondria and that it fulfills the function of the "redox response regulator" proposed by Allen (J. Theor. Biol. 165:609-631, 1993; Photosynth. Res. 36:95-102, 1993).
As a whole, the data obtained suggest that the oxidation state of glutathione is involved in the in vivo regulation of mitochondrial genome transcription in plants.
Financial support from the INTAS (Project
Number 97-0522) is acknowledged.
Return to the MNL 75 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}