Oregon State University
Coproporphyrinogen III oxidase is the eighth enzyme in the biosynthesis of tetrapyrroles in plants, catalyzing the oxidative decarboxylation of coproporphyrinogen III to protoporphyrinogen IX (Reinbothe, S. and Reinbothe, C. Plant Physiol 111:1-7, 1996). In plants, this and the earlier porphyrin biosynthetic steps are reported to take place exclusively in the plastid, where protoporphyrinogen IX can be converted to the first intermediate specific to chlorophyll biosynthesis or be processed towards the production of heme. Heme in the chloroplast serves as the precursor to the phytochrome chromophore, as well as providing for the synthesis of photosynthetic cytochromes. Protoporphyrinogen IX can also be exported from the plastid to the mitochondria where it is also converted into heme.
We cloned two genes encoding coproporphyrinogen III oxidase from maize. Using RFLP with recombinant inbreds, the genes were mapped to syntenous regions of chromosome 2S and 10L (Hardeman, K. et al., MNL 70:20-21. 1996). We have the entire genomic and cDNA sequence for Cpx1, which consists of 8 exons. For Cpx2, we are still missing the sequence for the last 3 exons. Sequence comparisons between the mature enzyme-encoding regions of the genes indicate a very strong preservation of identity, so that no portion of the proteins are less than 97% identical, including three highly invariant domains believed to be enzymatically critical. However, the N-terminal extension carrying the targeting information for each protein is unique and predicts targeting of the enzymes to different organelles.
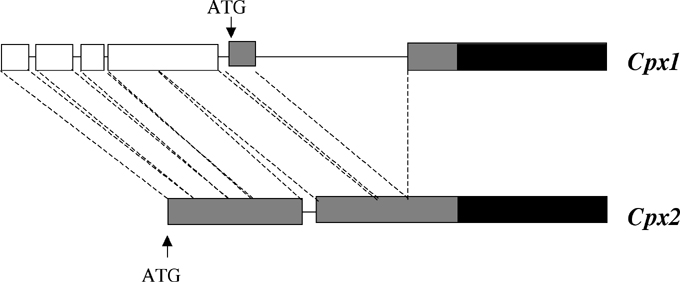
The Cpx1 gene encodes an N-terminal extension to the mature enzyme sequence that has the features of a chloroplast target peptide, as expected from studies in other plants (Madsen, O. et al., Plant Mol Biol 23:35-43, 1993; Kruse, E. et al., Planta 196:796-803, 1995). In Cpx2, however, 150 base pairs of this sequence are deleted, cleanly eliminating the transit peptide. Just 5' of this deletion, there is very substantial homology between the genes, but a series of small deletions/additions distinguish them. As shown in the figure, 5' UTR sequences of Cpx1 are homologous to an open-reading frame, headed by a methionine codon in Cpx2. This ORF is contiguous with the first exon, forming an N-terminal extension.
To test whether the N-terminal extension of CPX2 has potential targeting information, we applied a series of algorithms designed to look for and distinguish N-terminal peptides for chloroplast, mitochondrial and exported proteins. Using the PSORT, ChloroP, TargetP, and Predotar programs (Nakai, K. and Kanehisa, M. Genomics 14:897-911, 1992; Emanuelsson, O, et al., Protein Science 8:978-984, 1999; Emanuelsson, O. et al., JMB 300:1005-1016, 2000; Peeters et al., 1999, submitted), the CPX2 protein was strongly rejected as a chloroplast protein, and strongly predicted to be mitochondrial in location. To experimentally test this idea, we fused the putative Cpx2 N-terminal extension sequence to GFP to make a reporter gene driven by the 35S promoter, and introduced the construct into leaf epidermis biolistically. The GFP was found in small, dispersed spots, of a size and distribution that matched that seen when GFP was fused with a bona fide mitochondrial targeting peptide from CoxIV. We hypothesize, therefore, that a series of mutations has changed the targeting information and thereby the location of the coproporphyrinogen III oxidase encoded by Cpx2, but we have not yet demonstrated CPX activity in maize mitochondria.
In order to distinguish roles for Cpx1 and Cpx2, we looked at their expression patterns and we have begun to characterize the phenotypes of mutants in each gene. Using a semi-quantitative RT-PCR for detection, both Cpx mRNAs could be found in root, shoot, vegetative and reproductive tissue. The quantity of Cpx2 message did not vary much from tissue to tissue, and was at a similar level to Cpx1 in non-green tissue. However, Cpx1 mRNA was found at a level approximately three-fold higher than Cpx2 in leaf tissue. A Mu8-induced mutant in the Cpx1 gene was initially characterized in tandem with a very closely linked dek mutation (Hardeman, K. et al., MNL 1996). The dek seeds germinated into yellow seedlings whose leaves became necrotic, leading to plant death within about 2 weeks. This yellow phenotype is consistent with the function of CPX1 in the production of chlorophyll, and the necrotic phenotype is expected when blockage of the porphyrin pathway produces phototoxic tetrapyrrole intermediates. These phenotypes have also been reported for the Necrotic-4 locus (Hoisington, DA and Neuffer, MG, MNL 57: 159-160, 1983) which maps to the same small interval of chromosome 2. In a complementation test, one quarter of the cross progeny had a yellow, necrotic phenotype, indicating that Nec4 is probably the locus encoding coproporphyrinogen oxidase. In other plants examined, there is a single gene for coproporphyrinogen oxidase, and the enzyme activity is exclusively in the chloroplast. From the putative transit peptide, expression pattern, and mutant phenotype, we hypothesize that this is the role of the Cpx1 gene product.
Using the TUSC system in collaboration with Pioneer HiBred Inc., a line was found carrying a Mu insertion in the first exon of Cpx2 (cpx2-578). This mutant is viable and fertile as a homozygote, although we have not detected any normal mRNA produced from the cpx2-578 allele. The biological role associated with CPX2 is mysterious, but the strong conservation of the presumed active site sequences indicates that it encodes a working coproporphyrinogen oxidase, but of unknown function. It cannot compensate for a mutation in Cpx1. Perhaps earlier steps of tetrapyrrole biosynthesis occur in the maize mitochondrion than have been seen in other plants. Or, perhaps the Cpx2 coproporphyrinogen oxidase serves to detoxify superfluous tetrapyrroles in the mitochondrion. We are still crossing the extraneous Mu elements out of the cpx2 mutant line, so we are not yet sure whether there are milder phenotypes associated with it.
Figure:
Alignment of 5' UTR and exon 1 DNA sequences of the Cpx1 and Cpx2
genes. The boxes show homologous
DNA sequences and lines represent non-homologous sequences. Dashed lines
indicate the alignment of homologous DNA. Putative 5' UTR (white), translation
starts (ATG), code for the organellar targeting peptides (grey), code for
the N end of the mature enzyme (black).
Return to the MNL 75 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}