These proteins are induced by both cold and drought stress. Dehydrins (dlps), also referred to as Group II late embryogenesis abundant (LEA) proteins, are glycine-rich, hydrophilic, and thermostable. They have been hypothesized to function by stabilizing large-scale hydrophobic interactions such as membrane structures or hydrophobic patches of proteins (Close, Physiol Plant, 97:795-803, 1996). Dehydrins accumulate in response to cold in the nucleus or cytoplasm, but it is unknown if they can accrue in mitochondria or chloroplasts. In the previous study we have found the accumulation of two dehydrin-like proteins in the plant mitochondria after low temperature treatment (Borovskii et al., J. Plant Physiol., 156:797-800, 2000).
In this connection, the objective of this study was to determine whether dlps localize to maize mitochondria in response to stimuli other than cold adaptation treatment (10 C for maize).
Three-day-old etiolated seedlings of Zea mays (L.) (maize) were grown at 27 C. Unstressed plants were maintained under growth conditions for one day. Mild cold treatment (acclimation) was carried out by subjecting seedlings to a temperature of 10 C for 7 days. Freezing stress was performed at �10 C for 20-30 min until ice crystallized on the surface of seedlings. Transfer of two-day-old seedlings onto dry filter paper for 1 day at growth conditions served as a model of drought stress. ABA treatments were made at the control temperature by spraying 1mM ABA (Sigma) solution with 0.1% of Tween-20 (Sigma). ABA treated seedlings were harvested the day after treatment. Control and treated seedlings were compared at similar growth stages. Mitochondria were isolated according to the technique described by Borovskii et al. (2000).
Sonicated (Fig. 1) and unsonicated (Fig. 2) mitochondria were used further for extraction of total and thermostable proteins. Mitochondrial proteins of maize were fractionated by 10% SDS-PAGE (25 mg of protein per lane) using a mini-Protean PAGE cell (Bio-Rad) according to manufacturer's instruction. Western blotting and immunodetection were carried out as described by Timmons and Dunbar (1990). Antibodies against dehydrins were kindly provided by Dr. T. J. Close. All experiments were replicated three to four times.
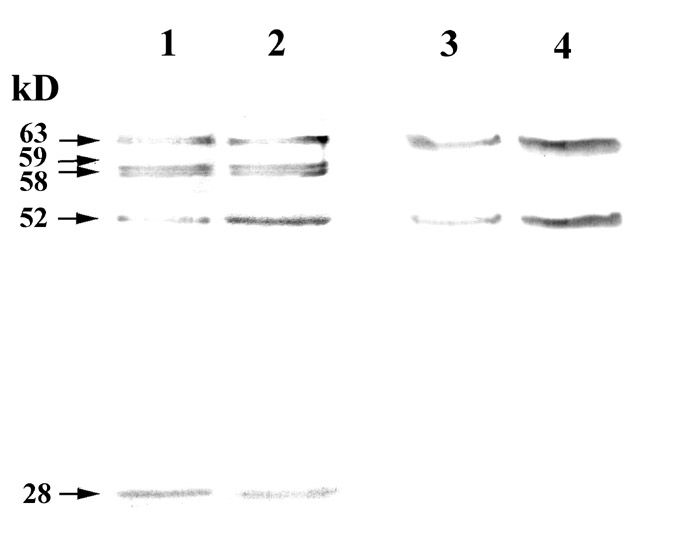
Five dlps were found in maize mitochondria (Fig. 1). Bands corresponding to all these proteins were very weak or absent when antibodies were blocked by dehydrin peptides (data not shown). Two of these polypeptides were thermostable, but the other three proteins seem not to be thermostable. Finding proteins immunologically related to dehydrins but constitutive and unstable to high temperature is unusual, but sometimes occurred.
Figure 1. Dehydrin-like proteins (dlps) of maize mitochondria after cold adaptation. Mitochondria from control (1,3) and cold-treated (2, 4) seedlings were disrupted by sonication. Total (1, 2) and thermostable (3,4) mitochondrial proteins were fractionated by 10% SDS-PAGE, electroblotted onto nitrocellulose membrane, and probed with antibody against dehydrin (1:1000). Molecular masses of dlps are indicated on the left.
The dlps with mol. masses 63, 52, 28 kD were found in the previous study (Borovskii et al., J. Plant Physiol., 156:797-800, 2000) in wheat and rye and apparently are "common" mitochondria proteins. Low temperature adaptation of maize resulted in the strong accumulation of "common" thermostable polypeptides with mol. masses 63 and 52 kD (Fig. 1) that was more pronounced in the thermostable fraction. Conversely, the unthermostable proteins did not accumulate. Apart from this, unthermostable dlps were not or very slightly induced by all the treatments used (Fig. 2). Based on this observation we concluded that these proteins were not involved in the stress reaction and adaptation.
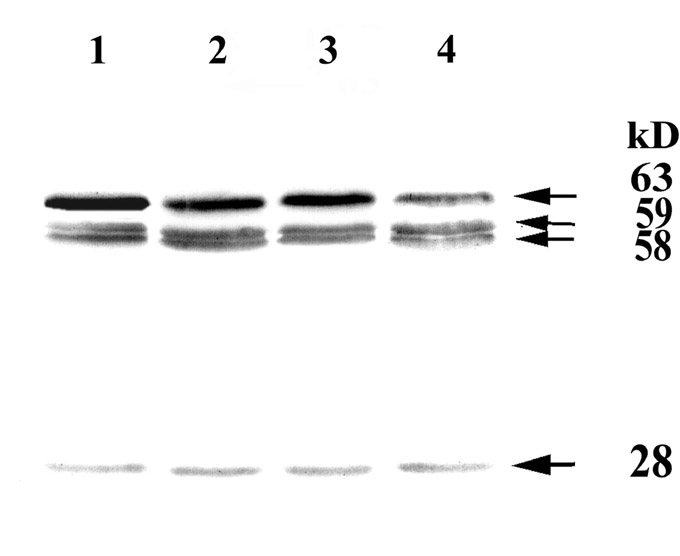
Figure 2. Mitochondrial dehydrin-like proteins from the control (1), freezing (2) and drought stressed (3) and ABA treated (4) maize seedlings. Protein from unsonicated mitochondria was subjected to separation and Western blotting (the same as in Fig. 1). Molecular masses of dlps are indicated.
The freezing and drought stresses had no effect on the accumulation of dlp 63 kD in mitochondria of maize (Fig. 2, ln. 2). The dlp52 was not observed under any treatment (Fig. 2). The differences in control samples of sonicated (Fig. 1) and unsonicated mitochondria (Fig. 2) illustrated to our minds that some groups of dlps had a strong association with large, slightly disruptive fragments of mitochondria. Such groups of proteins were discarded with undisrupted mitochondria. Because of that, dlp63 was stronger in Fig. 2 and weaker in Fig. 1 in the control samples.
Both proteins (dlp52 and dlp63) were not induced by ABA treatment in the maize mitochondria (Fig. 2, ln. 4). Moreover, ABA treatment resulted in decrease of dlp63. Apparently, augmentation of dlps concentration during cold adaptation is not associated with ABA accumulation, but dependent on low unfreezing temperature, as they slowly accrue during cold adaptation.
Thus, mitochondria of maize, a plant from a warm-temperature habitat, apparently slowly adapt to cold conditions and are unable to react quickly in severe stress conditions. It is likely to partially account for the chilling-sensitive nature of maize.
The research was funded by the Russian
Foundation of Basic Research (project 99-04-48121). We sincerely thank
Dr. T. J. Close for the gift of the dehydrin antibody and dehydrin-specific
peptide.
Return to the MNL 75 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}