The LMW HSPs range in size from approximately 17 to 30 kD and share a conserved C-terminal domain common to all eukaryotic LMW HSPs and to the a-crystallin proteins of the vertebrate eye lens. LMW HSPs function as chaperones preventing polypeptide damage. LMW HSPs form granular structures in cells, at the increased temperature associating with cell endoplasmic reticulum and plastid membranes, preventing their damage. In higher plants six nuclear gene families encoding LMW HSPs have been defined. Each gene family encodes proteins found in a distinct cellular compartment, including the cytosol, chloroplast, ER, and mitochondrion (Waters, E.R. et al., J. Exp. Bot. 47:325-338, 1996). Mitochondrial LMW HSPs (mit LMW HSPs) are nuclear-encoded stress-regulated HSPs, which play an important role in the protection of mitochondria and processes of oxidative phosphorylation. But the location of these LMW HSPs in mitochondria is still unclear. Methods of SDS-PAGE-electrophoresis and Western blot with antibodies to a-crystallin sequence were used to study localization of total and mitochondrial LMW HSPs in maize, wheat and rye cells. For comparison, we chose maize as a thermotolerant species, and wheat and rye as less tolerant species.
Three-day-old etiolated seedlings, grown at 27 C (maize) and at 23 C (wheat and rye) were subjected to heat shock (42 C, 3 h.) and used for the mitochondria isolation as described elsewhere (Borovskii, G.B. et al., J. Plant Physiol. 156:797-800, 2000). Isolated mitochondria were treated with pronase E (1 mg/ml) for one hour. Then mitochondrial proteins were extracted as previously described (Borovskii, G.B. et al., J. Plant Physiol. 156:797-800, 2000). Mitochondria without protease treatment were used for the extraction of the proteins of the "control".
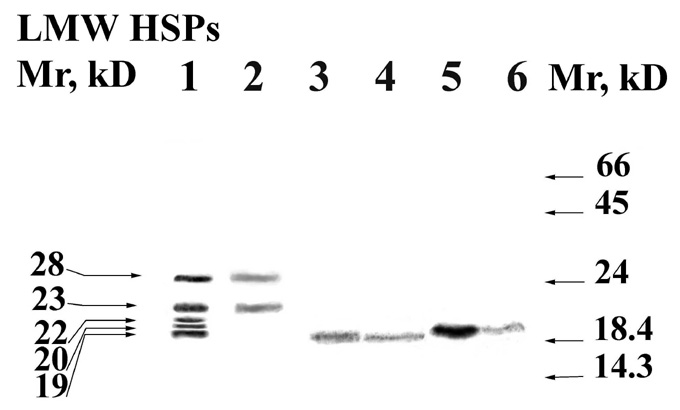
Western blotting demonstrated that LMW HSPs immuno-chemically related to a-crystallin appear among mitochondrial proteins of all the species under heat shock (Fig. 1). Five proteins were found in maize mitochondria and one protein was found in mitochondria of wheat and rye. Pronase treatment of mitochondria showed that maize mit LMW HSPs 22, 20 and 19 kD are located outside, but maize mit LMW HSPs 24 and 28 kD are located inside of the organelles. Apparently maize mit LMW HSPs 24 and 28 kD are matrix proteins. The absence of 29 and 23 kD in cytoplasm fraction supports this manner of location of LMW HSPs in maize mitochondria (Fig. 2). The location of maize mit LMW HSPs concurs with data of other authors, who identified LMW HSPs in soybean mitochondria by radiolabelling (Chou, M. et al., Plant Phys. 89:617-621, 1989). It was shown that shift of the seedlings to 28 C after heat shock leads to the appearance of organelle-associated LMW HSPs in cytosol. While the group of LMW HSPs 15-18 kD migrates from mitochondria, the 22 and 24 kD are always present in the mitochondrial fraction, which allows to submit their location inside of the organelles.
The treatment by pronase of mitochondria of wheat and rye with further protein extraction showed that HSP 20 kD are located outside and inside of organelles. In rye mitochondria the part of the 20 kD is localized inside that was stated by the decline of the coloration depth of the according spots. Other researchers confirm our results concerning wheat mit LMW HSP 20 kD (Basha, E.M. et al., Plant Sci. 141:93-103, 1999).
To determine the location of these LMW HSPs in other cell compartments, we extracted total and cytoplasm proteins as described above. Cytoplasm proteins were extracted from supernatant remaining after isolation of mitochondria and crude cell particles. It contained soluble cell proteins and proteins of membrane structures, nuclei and cell wall. The data showed that maize LMW HSPs 28 and 24 kD were specific for mitochondria, LMW HSP 20 kD of wheat and rye turned out to be not only mitochondrial but common cell protein (Fig. 2). The group of LMW HSPs 22-18 kD was found in the total, mitochondrial and cytoplasm protein fractions of maize.
The absence of two maize LMW HSPs 29 and 23 kD in the cytoplasm confirms the importance of these proteins for the mitochondria. Perhaps a�crystallin-related LMW HSPs attached to the mitochondria outside are able to migrate from the organelles to the cytosol after returning the cells to the normal conditions. Cognate process was discovered in soybean mitochondria (Chou et al., Plant Physiol. 89:617-621, 1989). Perhaps external LMW HSPs 20 and 22-18 kD are related to the class I LMW HSPs that originally localized in the cytoplasm. Material speaks well for this supposition about presence of extensive similarities between mit LMW HSPs and class VI LMW HSPs (Goping, I.S. et al., Plant Mol. Biol. 16:699-711 1991). The analysis of the primary polypeptide sequence for determination specific sectors for the VI class of LMW HSPs is required for verification of this supposition.
This work was supported by the Russian Fund of Basic Research (project 99-04-48121).
Figure 1. Mitochondrial proteins extracted from shocked seedlings (42 C, 3 h.) of maize (1, 2), wheat (3, 4) and rye (5, 6). Mitochondria were isolated from three-day-old etiolated seedlings and divided into two groups. One group of mitochondria was incubated with pronase E (2, 4, 6), the other one was used for reference (1, 3, 5). LMW HSPs immunochemically related to a-crystallin were identified. Molecular weight standards are indicated on the right. The relative molecular weights of LMW HSPs are given on the right.
Figure
2. Western blotting of total (1, 4, 7), mitochondrial (2, 5, 8) and
cytoplasmic (3, 6, 9) proteins extracted from shocked seedlings (42 C,
3 h.) of maize (1, 2, 3), wheat (4, 5, 6) and rye (7, 8, 9). Molecular
weight standards are indicated on the right. The relative molecular weights
of LMW HSPs are given on the right.
Return to the MNL 75 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}