Previous research indicated that the Myb domain is responsible for the DNA-binding activity (Grotewold et al., Cell 76: 543-553, 1994) and DNA-binding specificity (Grotewold et al., PNAS 97: 13579-13584, 2000) of P1. The P1-encoded protein contains an acidic region in the non-Myb C-terminal portion, between amino acids 207 and 242, suspected to serve as an activation domain (Grotewold et al., Proc. Natl. Acad. Sci. USA 88: 4587-4591, 1991). Chopra and co-workers (Plant Cell 8: 1149-1158, 1998) found that other alleles of P1 retain this region, providing further evidence of its significance for the P1-regulatory function. Although the P1-wr and P1-rr alleles differ in their C-terminal ends, the proteins that they encode contain identical Myb DNA-binding domains as well as identical acidic regions.
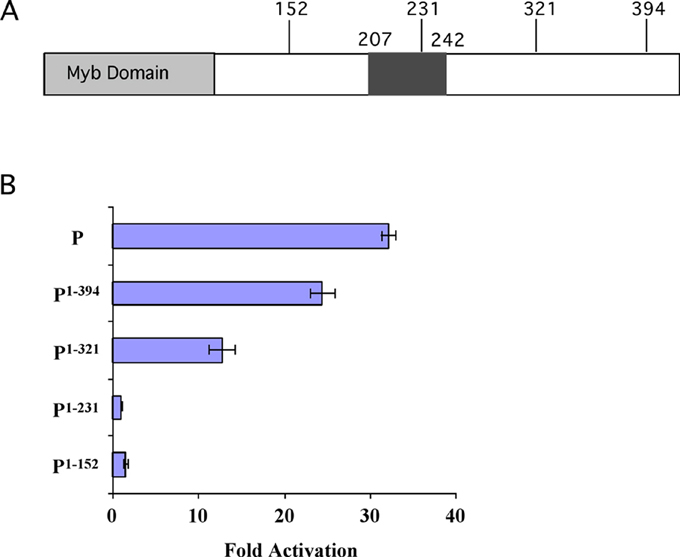
To more precisely define the regions in P1 important for its regulatory activity, truncations of the protein were generated. These truncations were cloned under a CaMV 35S promoter, and each truncation was then tested for its ability to activate a construct containing the A1 promoter driving the expression of the luciferase gene in transient expression assays as described (Grotewold et al. PNAS 97: 13579-13584, 2000). Figure 1A illustrates the position of the termination points of each construct. P1 provides the full-length protein, serving as a positive control. P1-152 excludes the putative activation domain, P1-231 contains approximately two thirds of it, and P1-321 and P1-394 contain the entire putative activation domain. As shown in Figure 1B, only those constructs which include the entire putative activation domain were able to activate transcription from the A1 promoter. P1-231 was not able to activate in spite of the fact that it contains about two thirds of the acidic region. These results suggest that residues in the region between amino acids 231 and 321 are essential for P1 activity, and that residues C-terminal to 321 can be deleted without a significant loss in P1 activity. Interestingly, this C-terminal part of P1 is the region that is different between the P1-rr and P1-wr alleles, which show distinct pigmentation patterns in the pericarp and cob tissues. Currently, experiments are underway to show whether the acidic region (207-242) is responsible for the essential nature of the 231-321 region.
Figure
1. (A) Various truncations made of the P1 gene, outside of the
Myb domain in the C-terminal region. Numbers indicate amino acid termination
points of each truncation. The putative activation domain spans from amino
acid 207 to 242 and it is represented as a black box. (B) Levels of activation
of the luciferase gene driven from the A1 promoter by P1
and the truncation constructs in transient expression experiments carried
out in BMS cells.
Return to the MNL 75 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}