University of Minnesota and USDA-ARS
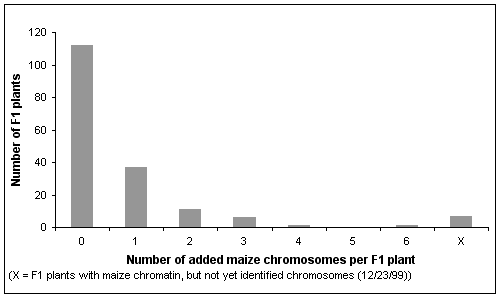
The principle for the production of oat-maize chromosome addition lines has been described previously (Riera-Lizarazu et al., TAG 93: 123-135, 1996). From more than 25,000 crosses of maize cv. Seneca 60 to the oat cvs. Starter, Gaf Park, Preakness, Kanota, Sun II, Stout, and an experimental hybrid (MN97201-1 x MN841801-1), we have rescued about 2000 embryos by in vitro culture on 1/2 MS medium. About 12 % of the F1 embryos germinated, and 175 of them grew to vigorous plantlets which were analyzed by molecular and cytogenetic means. The majority of these F1s were haploid oat plants without maize chromosomes. Approximately 36 % of these F1s were partial hybrids, however, retaining one or more maize chromosomes (Figure 1). We observed a maximum of six retained maize chromosomes in addition to the haploid oat complement in one plant. Most plants had lower numbers of added maize chromosomes (Figure 1). The generally low numbers of retained maize chromosomes among the recovered F1 hybrids indicate a competitive advantage for cells having eliminated maize chromosomes. The specific chromosomes and their allelic constitutions lead to distinct phenotypes.
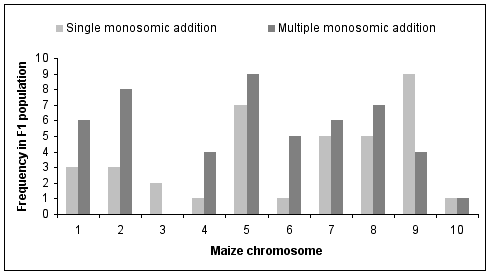
Figure 2 summarizes the identity of the added maize chromosomes and their frequencies in the F1 hybrid population. A total of 87 maize chromosome additions have been identified to date. In 37 F1 plants a single maize chromosome was retained along with the haploid oat complement. All ten maize chromosomes have been recovered as single monosomic additions. Maize chromosome 9 was most frequent among the single monosomic additions. Chromosome number 5 was slightly less frequent as a single addition, but most frequent in combination with other chromosomes. Apart from maize chromosome number 3, all of the other chromosomes also were found together in combination with other chromosomes added to the haploid oat complement. There was no obvious preferential combination of the maize chromosomes among the multiple additions. The double monosomic additions involved the combinations of the maize chromosomes 5 + 9, 5 + 8, 1 + 5, 2 + 6, 1 + 8, 2 + 5, 4 + 7, 5 + 6, 4 + 6, and twice 2 + 7. Triple monosomic additions included the chromosome combinations 6 + 7 + 9, 2 + 7 + 8, 2 + 6 + 9, 2 + 5 + 8, 1 + 4 + 5, and 1 + 5 + 8. In one F1 plant we identified the added maize chromosomes 1 + 2 + 8 + 9. And one F1 plant had the maize chromosomes 1 + 4 + 5 + 7 + 8 + 10. The most important result is that we detected for the first time an entire set of oat-maize chromosome addition plants, which carry singly each of the ten maize chromosomes.
Haploid oats are known to produce up to 40 % seed set (Davis, MS Thesis, University of Minnesota, 1992). As it occasionally occurs with haploids of other allopolyploid and amphidiploid cereals, e.g. wheat and triticale, haploid oats can produce doubled haploid or even aneuploid (e.g. monosomic or nullisomic) offspring by completely or partially unreduced gametes. The high seedset frequency in oats, however, is mainly caused by restitution of meiosis I. Meiotic restitution results in unreduced spores, which eventually generate fertile gametes. Our haploid F1 hybrids with or without maize chromosome additions occasionally underwent the same or very similar processes. As a result, hexaploid oat plants with nullisomic, monosomic, or disomic additions of maize chromosomes were formed among the F2 genotypes. We recovered disomic additions for maize chromosomes 2, 3, 4, 6, 7 and 9 and a monosomic addition for maize chromosome 8. Details on this material and its availability are presented in Table 1.
To produce radiation hybrid lines a disomic chromosome 9 addition line was backcrossed to oat to generate a line monosomic for maize chromosome 9. These seed were irradiated, grown to maturity, and allowed to self pollinate. Seed from individual panicles were planted and tested for the presence of maize DNA. Approximately 10% of the progeny carried maize DNA. Lines carrying maize DNA were characterized cytologically and with molecular markers. We have made over 40 radiation hybrid lines for chromosome 9, and are developing radiation hybrid lines for chromosomes 2 and 4.
At present we are working with 38 radiation hybrid lines for chromosome 9. Our first step has been to map markers previously mapped to either the consensus SSR map or the 1998 UMC map. This allows us to look at the distribution of breaks along chromosome 9 and provides a basis for future comparisons between the radiation hybrid map and other chromosome 9 maps. The first 56 markers placed on the map defined 41 segments. Many more markers need to be mapped before we can be confident that the chromosome breaks are adequately defined.
A major effort this year has been to develop new markers. Using EST sequences from ZmDB (http://www.zmdb.iastate.edu/), primers have been designed and tested. To date 385 primers were developed that amplified maize DNA and distinguished maize from oat sequences. These primers have been mapped to chromosome. Using our new oat-maize addition lines a total of 178 EST sequences have been allocated to chromosome 2 (27 ESTs), chromosome 3 (34 ESTs), chromosome 4 (46 ESTs), chromosome 6 (28 ESTs), and chromosome 9 (23 ESTs). Thirty-four EST markers were located to chromosome 9; 11 of these markers were also located on one or more of the other chromosomes tested. These markers are being placed onto the chromosome 9 radiation hybrid panel. 20 ESTs showed loci on more than one chromosome tested. One EST sequence, AI737657, mapped at the Sh1 locus. A BLAST search revealed that this EST was a Sh1 sequence.
The data for 385 EST primer sequences, their chromosome allocation and the description of the radiation hybrid lines will soon be available on our website: http://www.agro.agri.umn.edu/rp/genome/
This material is based upon work supported by the National Science Foundation under Grant No. 9872650.
Figure 1: Frequencies of the number of maize chromosomes retained in 175 oat- maize F1 plants.
Figure 2: Frequencies of 87 identified maize chromosomes as single and multiple monosomic additions in 56 oat-maize F1 plants.
Table 1. Oat-maize single monosomic
and disomic addition lines.
|
|
Availability | |||||
| Added maize chromosome | Oat background | Before 1999 | In 1999 | Offspring
F2, F3, F4, ... |
Seed | DNA |
| 1 | Preakness
Starter Gaf Park |
1 | 1
2 |
0 | None
None None |
Plenty
Limited Limited |
| 2 | Starter | 4 | 3 | > 100 | Plenty | Plenty |
| 3 | Sun II
Preakness Starter |
1 | 1
1 |
> 100
29 0 |
Plenty
None None |
Plenty
Limited Limited |
| 4 | Starter | 5 | 1 | > 100 | Plenty | Plenty |
| 5 | Starter
Gaf Park Hybrid |
1? | 3
2 2 |
0 | None
None None |
Limited
Limited Limited |
| 6 | Starter
Dumont |
1 | 1 | > 100
> 100 |
Plenty
Plenty |
Plenty
Plenty |
| 7 | Gaf Park
Starter |
1 | 5 | > 100
> 100 |
Plenty
Plenty |
Plenty
Plenty |
| 8 | Gaf Park
Starter |
1
1? |
2
3 |
> 100* | None
None |
Limited
Limited |
| 9 | Starter
Kanota Gaf Park |
2 | 6
1 2 |
> 100
1 30 |
Plenty
None None |
Plenty
Limited Limited |
| 10 | Gaf Park | 1 | None | Limited | ||
* monosomic offspring only
Return to the MNL 74 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}