North Carolina State University
John Doebley planted seed of the Hruska collection in a growth chamber, observing:
"Plants produced mature seed four months after planting, this much more rapidly than is typical of this species [Zea luxurians]. This collection, clearly differentiated from the Guatemalan form of this species by its small stature and early maturity, may represent a new subspecies." (label on specimen Doebley No. 648).
I saw plants of the CIMMYT accession being grown for seed replication in the field station at Tlaltizapán, Morelos (940 masl), in early 1995 (planted in July, 1994). They were profusely tillered, standing about 1.6 m tall, and were flowering. I asked Dr. Suketoshi Taba to bring a plant to Texcoco (near Mexico City, 2255 m) after the seed were harvested, on the suspicion that it would continue growing. This done, the plant was cut into three parts, each of which grew well, one flowering twice more. The 1994 clones at El Batán were preparing to flower at the end of 1996 when killed by frost.
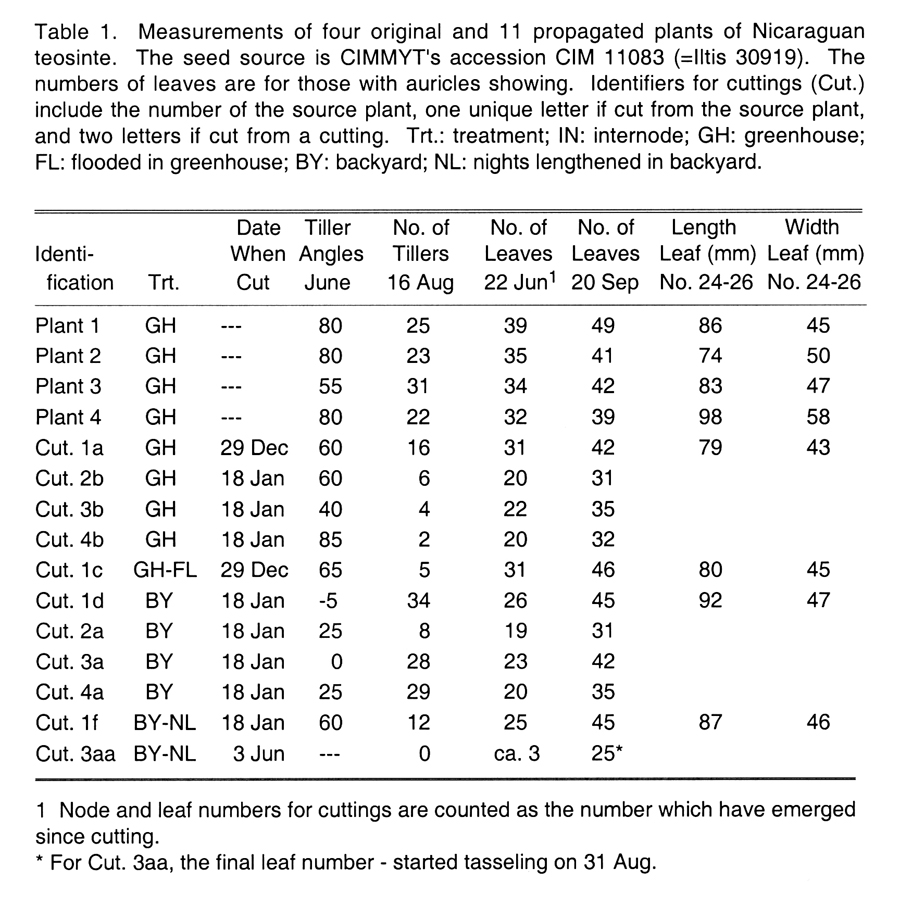
Shoot morphology: After soaking the fruit cases (rachis segments) in diluted pharmacy H2O2 (20%) for over an hour, I planted original and self-pollinated seed of CIM 11083 in a screenhouse at CIMMYT's El Batán station in March, 1996, and, on 25 Sep 98, in a North Carolina State University greenhouse in Raleigh, NC. For months the two stands grew similarly, but there was segregation of some interesting traits (Table 1). In Raleigh, the four plants started growing tillers four weeks after planting, at the lowest nodes of the main stem, and then at the lower nodes of larger primary tillers. Tillers eventually totaled 22-31, up to 15 being primary (on the main stem).
A week later, starting a new phase of juvenile Stage 1, about half the plants were prostrating themselves on the ground (for Plant 1, main stem at 20° from the ground, tillers at 0°), growing like maize with the lazy gene, while the rest relaxed to 50°-70°. By this time there were 5 leaves with auricles showing. At this point, I commenced numbering every fifth leaf on the main stem and the largest tiller.
At the start of Stage 2, 15 weeks after planting (8 Jan 99), the stems of all plants were growing more upright, including the 15-28 tillers. Main stems had 17-21 leaves. The main stems were inclined at 30°-80°, tillers at 40°-60°. A few more tillers were produced before 14 Feb, none thereafter. Perhaps the pots were too small, although fertilizer was added occasionally. Stage 2 continued into the autumn for many plants -- lights in nearby greenhouses and then long days prevented flowering. However, in cuttings transplanted to a garden, flowering was induced by artificially lengthening nights with black plastic covers (below).
As plants have matured, removal of old leaf sheaths has revealed very short internodes and masses of secondary prop roots. The approximate distances from node 1 to node 20 of the main stems of the original four plants are 3.5 cm, 7.5 cm, 11.5 cm and 10.2 cm. I recommend measuring internodes as soon as possible, before the secondary roots grow. Average internode lengths in the interval between nodes 20 and 25 of the four plants are 1.0 cm, 1.4 cm, 1.2 cm and 1.7 cm (the two shorter are the two more prostrate plants in Stage 1). Higher internodes are longer.
It has been easy to cut tillers and root them. Three were taken from Plant 1 on 29 Dec 98, and three were cut from each of the 4 original plants on 18 Jan 99. Those that were about 1 cm in diameter did well, better if a bit of root had emerged. The propagated plants grew much like the originals, but those cut after December did not recline as much, UNLESS they were transplanted outdoors (16 Apr) where angles were 40° to 60° less than the equivalents indoors (Table 1). The three cut in December from Plant 1 were lying flat or nearly so (0°-20°) by 9 Feb, and remained nearly prostrate through 21 Apr, even though they were treated in varying ways (below)! Those of Plant 3 reclined at an intermediate angle. Two cuttings made on 3 June have not tillered. Perhaps cuttings were imprinted by the stage they were in when cut, but crowding of plants, root binding and temperature seem to have had major effects. One January cutting of each original was transplanted to my backyard on 16 Apr, where those of Plants 1, 3 and 4 eventually produced 28-34 tillers and enormous plants; the Plant 2 cutting has remained smaller, eventually becoming shaded by the others.
Leaf counts show that cuttings reflect their source plants in relative numbers of leaves grown per month; Plants 2 and 4 had fewer leaves by 22 June than Plants 1 and 3. New leaves emerged in intervals as short as four days.
Leaves of this teosinte bear small (0.4 mm long) hairs visible with a loupe, but only on the upper surface of the blade, not the sheath or auricle. Leaf blade margins have sharp, apically directed barbs 0.06 mm long which can cut skin. When sheaths are rubbed upward, one feels (especially with the lips) a slight, downwardly directed roughness. Shapes of blades will be discussed.
A flooding experiment: Because of this ability to replicate plants of Nicaraguan teosinte, it has been possible to run several experiments on the "same" plant. Cuttings of Plant 1 ended up: (a) in a tank of water, (b) in 15 cm and 25 cm pots in the greenhouse, and in my backyard, (c) with and (d) without night-lengthening.
Plants in Chinandega, Nicaragua had been observed growing up to 5 m tall, yet none of the progeny grown in Mexico grew over 2 m tall, so I have been testing a hypothesis that extended flooding of the plants greatly stimulates growth. On 16 Feb one of the December cuttings (Plant 1, Cut. 1c) was placed in a tank of water so that the roots, but not the shoots, were constantly underwater (another plant was submerged 5 cm deeper but suffered badly so after 16 days it was removed). Cut. 1c did not recline as much as twin cuttings. It has grown well in a 25 cm pot. By 5 June it had grown only 7 tillers, while an unflooded twin had 12. On 23 Sep it was growing taller than the controls and other plants, about 1.8 m to the tallest auricles, versus 1.0-1.3 m. It, as the others experiencing short nights, remains in vegetative Stage 2. It continues to grow ever taller than the others.
Cuttings of Plants 2 and 3 (Cut. 2c & 3c), in 15 cm pots, were placed in a separate water tank on 22 Apr, with the roots just under water. These two were lowered 6 cm on 5 June, and even more about a month later. One soon rotted at the base and the other later, but nodal roots have grown and the plants still live, hydroponically. All three plants have grown masses of secondary roots in the water, with small tertiary roots 0.2-0.4 mm in diameter extending upward, either 1/2 cm above the soil surface, or 1.5-2.5 cm towards the water surface. The small roots are dense, about 40 per 3 cm of secondary root. A bluish green alga has been found growing over the roots of the cutting from Plant 1, possibly affecting its growth.
Photoperiod adjustment: In North Carolina, short summer nights and extraneous greenhouse light in the winter has prevented flowering, so artificial night-lengthening was applied to cuttings of Plants 1 (cut on 18 Jan) and 3 (cut on 3 June). Three of an additional four plants grown from seed from Nicaraguan teosinte plants pollinated in Mexico (planted 16 Apr 99) were also treated, as were plants of Z. luxurians and of Z. mays ssp. huehuetenangensis. Frames 4 ft. high were covered in black plastic, 4 mil thick. These were placed over the plants for 21 nights (17 July-7 Aug), 13 hours each night. By 13 Aug it was obvious that changes had occurred -- emerging leaves had a different shape (below).
A tassel appeared on Cutting 1f on 19 Aug, silking started on 29 Aug and pollen shed started on 31 Aug. Tassels on Cutting 3aa appeared 6 to 16 days later, but shed much later, slowed by cool weather after 16 Sep or by crowding. Perhaps the older the plant or cutting, the more quickly it can be induced to flower, or being a cutting from a cutting may have affected the later one. Seedlings may be ready to flower later than cuttings made on the planting date. None of the untreated plants showed a sign of flowering by 22 Sep, but by 25 Oct branches were growing from upper nodes and leaves were broadening at the base.
Flowering stage: The third, reproductive stage of shoot development is quite different. The forms of the tassels, ear branches and earlets will be described in a later report. As reproductive structures developed, upright stems, leaves and upper branches changed. An indicator of Stage 3 was the shape of the leaves, broadened at the base and shortened. Broadening started at nodes roughly where the lowest ear branches occurred. Widths at 5 cm from the ligule, for leaves in the middle of the zone of ear branches, ranged 45-59 mm, lengths ranged 54-70 cm, and maximum widths ranged 49-63 mm at 30-45% of the distance from ligule to tip. Grown outdoors, leaves on untreated plants, still in Stage 2, reached 1.4 m long. Their widths at 5 cm from the ligule were only 15-30 mm. Their maximum width was at more than 1/2 of the ligule-tip distance. On plants at Stage 2 in the greenhouse, leaves were smaller in all dimensions but had the same shape as the outdoor vegetative cuttings.
Discussion: Several trends and much variation have been noted. While taking measurements and notes, many questions occurred to me. Maybe the flooding noted at the Apacunca site ebbs and flows with the size of storms. Would the plants grow taller and better if flooding alternately covered stems for up to 30 cm and uncovered roots to a depth of 5 cm? Might the 5 m height of some plants at the collection site be due to introgression from maize? F1 hybrids of maize with Z. luxurians and this teosinte reach that height. What are the youngest age, shortest days and fewest days of treatment which will give consistently good flowering? Does the "quality" of a cutting change over time or over generations of cutting? Do cool temperatures increase the "lazy" effect? In many ways this is a most unusual teosinte.
Return to the MNL 74 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}