University of Northern Iowa
Emporia*, Kansas
Department of Biological Sciences
Although the origin of the maize ear is still not unequivocally known, researchers investigating teosinte inflorescence development have revealed a developmental pattern common to all Zea (maize and teosinte) inflorescences (Sundberg and Orr, Amer,. J. Bot. 77: 141-152, 1990). However, key developmental features have been uncovered that differentiate ears from tassels and teosinte inflorescences from maize inflorescences (cf. Fig. 25, Orr and Sundberg, Amer. J. Bot. 81: 598-608, 1994). It is especially interesting that one observation in these investigations disclosed a key organogenic event to account for the evolution (distichy to polystichy) of a multiple-ranked maize inflorescence from a two-ranked inflorescence. A key trait in the evolution of the maize inflorescence, acquisition of polystichy, may have arisen from a change in a developmental program that regulates the spikelet pair primordia condition. Spikelet pair primordia in teosinte and maize give rise to two spikelet primordia only. In the "standard exotic" maize (Argentine popcorn) two inflorescence phenotypes were observed: intermediate distichy/polystichy (two-ranked and four-ranked), and wild type four-ranked polystichous inflorescences (Sundberg, LaFargue and Orr, Amer. J. Bot. 82: 64-74, 1995). In the intermediate phenotype the distichy condition was distal to the polystichy condition: the proximal portion of intermediate inflorescences bore eight rows of spikelets in four ranks, and the distal segments produced four rows of spikelets in two ranks. In wild type teosinte and maize inflorescences the apical meristem first produces spikelet pair primordia, each of which bifurcates to produce paired spikelet primordia. In the intermediate inflorescences of Argentine Popcorn a second cycle of bifurcation facilitates a switch from a two-ranked (distichy) condition to a four-ranked (polystichy) condition. Perhaps the origin of polystichy in maize was derived from a change in developmental regulation of meristem determinacy (Sundberg, et al., 1995). Regulation of maize inflorescence meristems that produce an additional number of meristems has been reported (Orr, Haas and Sundberg, Amer. J. Bot. 84: 723-734, 1997; Chuck, Meeley and Hake, Genes Dev. 12: 1145-1154, 1998; Lenhard and Laux, Cur. Opin. Plant Bio. 2: 44-50, 1999).
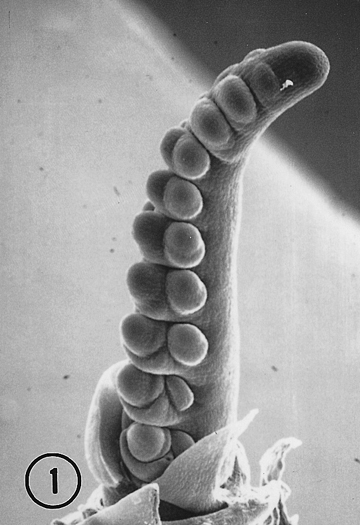
We used scanning electron microscopy to characterize the organogenesis of Toluca teosinte inflorescence, and to compare the development to other Zea inflorescences. Seed from the CIMMYT Maize Germplasm Bank was kindly given to us by Sukitoshi Taba. Plantings were made in a growth chamber under a long day, 15:9 hr light/dark cycle, and at the V5 (fifth vegetative leaf) stage plants were shifted to 8:16 hr light/dark cycle. Congruent with the light/dark cycle, the temperature was maintained at 24:18 C. Light intensity was maintained at 600-700 uE m-2 sec-1 (PAR). Our SEM examination of approximately 35 developing ears confirmed that in most ears (33/35) spikelet pair primordia were produced in two ranks (Fig. 1). However, rather than the expected pure distichy pattern in tassels most (25/30) spikelet pair primordia were produced in either three or four ranks (Fig. 2). As we expected, Toluca spikelet organogenesis in tassels and ears followed a pattern previously described in other teosintes. Each spikelet produced an upper and lower floret primordia, and each floret primordia produced three stamens and a gynoecium. Femaleness in Toluca spikelets was marked by the abortion of the lower florets and the abortion of the stamens in the upper florets. Male Toluca spikelets were marked by a retention of the lower florets, and an abortion of the gynoecium in both the upper and lower florets. This supports the hypothesis that both femaleness and maleness in teosintes are derived from and expressed on a common background (cf. Fig. 25, Orr and Sundberg, 1994). However, unlike the paired spikelet condition in Toluca tassels, Toluca ears displayed solitary sessile spikelets derived from arrested growth and abortion of pedicellate spikelets (Fig. 1). The abortion of pedicellate spikelets in female inflorescences was observed in other teosinte ears (Orr and Sundberg, 1994). This is further evidence that this morphological marker is sufficient to discriminate, at a very young developmental stage, teosinte inflorescences sexually and teosinte ears from maize ears.
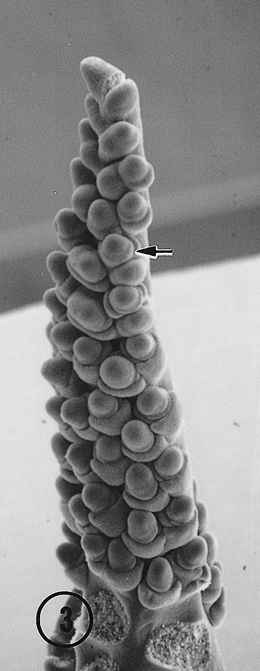
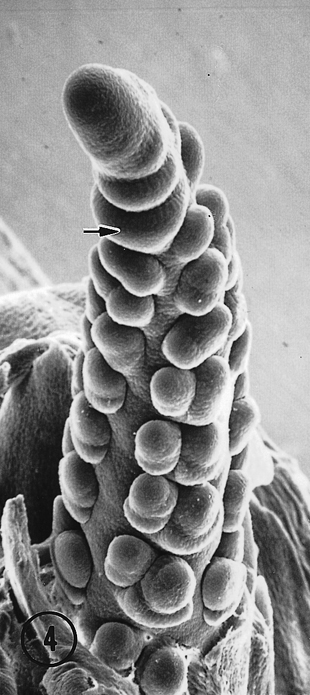
The most striking observation in this study of Toluca inflorescences was the occurrence of intermediate (distichous and polystichous) and pure polystichous tassels, and intermediate ears. In the population of 30 tassels we examined ten were intermediate, and 15 were 100 percent, four-ranked polystichous (Fig. 2). In the intermediate state, tassels (Fig. 3) and ears (Fig. 4) exhibited the distichy condition at the distal end of the inflorescence. This morphological intermediate phenotype is similar to that noted above for maize Argentine Popcorn (Sundberg, et al., 1995). In female inflorescences no purely polystichous ears were observed, and only two, three-ranked intermediate ears (Fig. 4) were noticed in the population of 35 we surveyed. It remains unclear why the frequency of additional ranks in the inflorescences of the Toluca population is much lower in the ear than the tassel. Also, it is obscure why an inflorescence meristem that initially produced three or four ranks shifted to make two ranks. We are currently examining additional Toluca inflorescences for evidence of the polystichous condition.
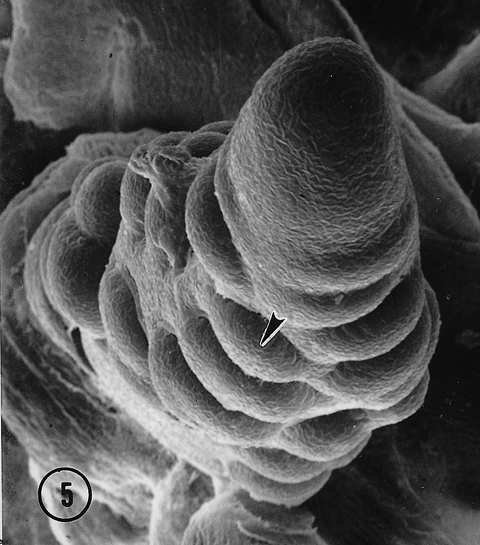
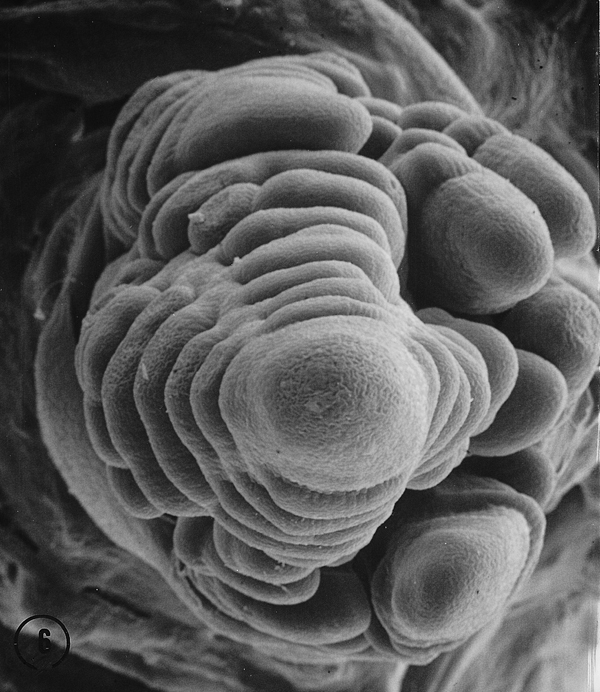
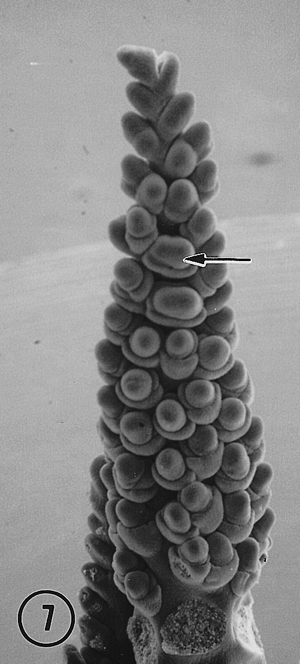
In each of the intermediate ears the distal portion of the inflorescence was distichous with two ranks of lateral primordia. In one ear a third rank arose at the fifth node from a division of a spikelet pair primordium at the fourth node (Fig. 4, arrow), suggesting that a second bifurcation of the original meristem resulted in two ranks of spikelet pair primordia. Thus, one rank with a potential for two rows became two ranks that resulted in four rows. A three-ranked, six rowed female inflorescence resulted. There is some evidence suggesting an additional bifurcation of each original spikelet pair primordia probably was a key event in the switch from distichy to polystichy in the evolution of a maize ear from a teosinte ear (Sundberg, et al., 1995). The other ear that exhibited an intermediate condition produced a partial third rank de novo on the adaxial surface of the inflorescence (Fig. 5). Intermediate and polystichous tassels exhibited both three and four ranks. Pure polystichous, eight rowed tassels arose from the formation of four ranks of spikelet pair primordia arranged in a decussate pattern (Fig. 6). A six rowed tassel or ear produced only three ranks of spikelet pair primordia with primordial ranks shifted toward the abaxial side. Additional rows (ranks?) in the proximal region of intermediate tassels appear to arise from a second bifurcation of lateral primordia (Fig. 3, arrow), or were derived from a de novo event (Fig. 7, arrow). It is also apparent in Fig. 7 that these de novo primordia divide into rows of paired spikelets. It would appear that these de novo primordia function as spikelet pair primordia. Quite possibly, but speculatively, the occurrence of these natural intermediate and pure polystichous inflorescences were the result of introgression between maize and Toluca teosinte. If this view is correct, this is the first developmental inflorescence study of a natural maize-teosinte hybrid and thus, may offer insight into the effect(s) on inflorescence development when different developmental programs are recombined in maize-teosinte hybrids. Interestingly, the first backcross of maize following F1 maize x teosinte hybridization resulted in polystichous ears (see Fig. 11B, Wilkes, Econ. Bot. 31: 254-293, 1977).
Finally, our studies provide evidence that unique phenotypes may arise in a hybrid population. In addition to the de novo spikelet pair primordium we noted above, we also found some tassels with three spikelets, rather than the usual two found in teosinte and maize. A similar occurrence of the latter was noted in other studies of inflorescence development: F3 plants derived from a cross of Race Reventador maize by Z. Mays subsp. parviglumis teosinte (see fig. 5, Doebley, Exp. Mol. Approch. to Plant Syst., Mo. Bot. Gard. 53: 57-70, 1995); the maize mutant Fascicled ear (Orr, Haas and Sundberg, 1997); the primitive maize Chapalote (Sundberg and Orr, Amer. J. Bot. 83: 1255-1256, 1996); and the primitive maize Argentine Popcorn (Sundberg, et al., 1995).
Fig. 1. SEM of developing Toluca teosinte ear with two (distichous) ranks. One rank is hidden behind the central axis. Spikelet pair primordia have divided into paired spikelets. At the base of the right row note the reduction in growth (and eventual abortion) of the pedicellate spikelet.
Fig. 2. SEM of developing Toluca tassel with four (polystichous) ranks of spikelet pair primordia. One rank is hidden behind the axis.
Fig. 3. An intermediate Toluca tassel with four ranks (eight rows) of spikelets on the lower two-thirds of the central axis, and two distal ranks of primordia on the upper third of the inflorescence. Two ranks (four rows) are partially hidden behind the axis. Basal tassel branches were removed to fully reveal the central axis. Proximal spikelets are characterized by outer and inner glumes. Note the basal spikelets include an outer lemma. Additional rows (ranks?) appear to arise from a second division of a lateral spikelet pair primordia (arrow).
Fig. 4. An intermediate Toluca ear with three ranks. The plane of distichy at the distal end is approximately at a right angle to the photo. A third rank is somewhat visible along the back, right edge of the axis. The central rank appears to arise from a division of a spikelet pair primordium. An older proximal primordium has undergone a second division resulting in two rows of spikelets.
Fig. 5. A polar view of a developing intermediate Toluca ear with three ranks of spikelet pair primordia. Note the apparent de novo production of a third rank (arrowhead).
Fig. 6. A polar view of a pure polystichous Toluca tassel with four ranks of spikelet pair primordia along the central axis. A bifurcation of these primordia is seen at the proximal end of the inflorescence. Basal lateral branches with a distichous arrangement of spikelet pair primordia are apparent
Fig.
7. A central spike of an intermediate Toluca tassel. Basal tassel branches
were removed to fully reveal the central axis. This specimen shows an apparent
de novo formation of an additional rank of spikelet pair primordia (arrow).
Note the bifurcation of this additional rank of spikelet pair primordia
in the formation of paired spikelets.
Return to the MNL 74 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}