Wuhan University
Beijing, China
Chinese Academy of Sciences
Guo et al.(1997 and 1998) crossed maize inbred line Zi 330 with DP, and made backcrosses of the hybrids with Zi330. Parthenogenesis was successfully induced in the BC1 by chemicals and the alloplasmic pure line 540 was obtained at the second generation of chemical-induced parthenocarpy (Pa2)(Guo et al. 1998). The pure line 540 was proliferated by inbreeding and crossed with maize inbred line Zi 50003. The F1 (the first hybrid generation) named Yi Dan 6 has already been applied to maize production in China. Both 540 and the F1 hybrids, Yi Dan 6, showed much stronger Helminthosporium turcicum and Helminthosporium maydis resistance and wider adaptation to stressful conditions, compared with the maize parents and the F1 intraspecific hybrids of maize (Guo et al.1997). It could be deduced that the DP chromatin dictating these characters had been introgressed into the parthenocarpic progenies (Pa2), the 540 should be an alloplasmic pure line and it had transferred the introgressed chromatin to its F1 hybrids, Yi Dan 6.
The karyotypes and chromosome C-banding patterns among maize Zi330, DP and the alloplasmic pure line 540 have already been analyzed comparatively (Yang et al., 1995; Yan et al., 1997). Although some chromosome variations of 540 were found in their studies, it was still unknown whether the DP chromatin was introgressed into the parthenocarpic progenies or not and if it was, where it was located. GISH using total genomic DNA as a probe to identify alien chromosomes or chromosome segments was well-established in many other plant species (Le et al. 1989; Heslop-Harrison et al 1990; Leitch et al. 1990; Mukai and Gill 1991; Schwarzacher, et al. 1992; Anamthawat-Jonsson and Heslop-Harrison 1993.; Kenton, et al. 1993; Mukai and. Nakahara 1993; Yasuhiko and Yumiko 1993; Chen and Armstrong 1994; Jellen et al. 1994; Jiang and Gill 1994; King et al. 1994; Fernandez-Calvin et al. 1995; Humphreys et al. 1995; Jacobsen, et al. 1995; Leggett and Markhand 1995; Keller et al. 1996; Zhong et al. 1996; Garriga-Caldere et al. 1997; Pickeing et al. 1997; Tang et al. 1997; Belyayev and Raskina 1998; Chen et al. 1998; Mikhailova et al. 1998; Stevenson and Armstrong 1998; Kamstra et al. 1999), while, as we know, no report about this respect has been published yet in the distant hybrids of crosses of maize with DP. In this study, the introgressed DP chromatins were detected and located in both the pure line 540 and Yi Dan 6 by genomic in situ hybridization.
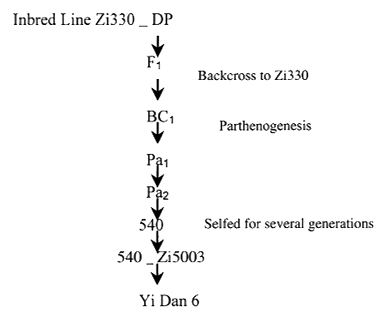
Zea diploperennis (DP) was supplied by Mr. Li Dongyu, Guangxi Zang Autonomous Region of China. Maize Inbred lines Zi330, alloplasmic pure line 540 and the F1 hybrids of 540 X Zi 50003, Yi Dan 6, were supplied by Institute of Genetics, Chinese Academy of Sciences, Beijing. Their pedigree is given in Fig. 1.
Genomic DNA was extracted from DP and inbred line 330 following the procedure described by Doyle and Doyle (1990). The total genomic DNA from DP for use as labeled probe was mechanically broken to 10-12kb fragments prior to labeling, and the total genomic DNA from 330 was fragmented to pieces about 100-250bp by autoclaving for 5min and used as blocking DNA according to the procedures reported by Heslop-Harrison et al. (1990), Schwarzacher et al. (1992), and Jacobsen et al. (1995).
Figure 1. Brief illustration of the relationships between the tested materials.
The preparation technique was that described by Song et al. (1994) with some modification. The harvested root tips were pretreated in saturated a-bromonaphthalene solution for 2hr at room temperature (RT). After rinsing in tap water, the root tips were fixed in fresh fixative (3 methanol: 1 glacial acetic acid) for 2-3hr at RT and then, digested in 1% cellulase and 1% pectinase (SERVA) at 28 C for about 2.5hr. The softened tips were subsequently rinsed gently with distilled water. Following removal of the water, 3-4 root tips were smeared with fixative onto a clean slide. The slide was flame-dried.
Nick translation of DP genomic DNA was adopted with the kit offered by Sino-American Biotechnology Company, China. 15µl of dNTP (mixture of average amount of dATP, dCTP and dGTP), 5µl of 10 buffer, 4µl of DP genomic DNA, 5µl of Bio-11-dUTP and 5µl of DNAse1 and DNA polymerase1 mixture, and ddH2O were added to total volume 50µl. It was incubated at 15 C for 2hr, and the reaction was stopped by adding 5µl of 0.2M EDTA. The labeled probe was purified by passing the reaction solution through a Sepharose CL-6B (Sigma) column.
The procedures of in situ hybridization and signal detection followed the modified methods published by Leitch and Heslop-Harrison (1993), Schwarzacher et al. (1992), Schwarzacher (1994) and Jacobsen et al. (1995). The hybridization mix was 42µl in the total volume and it contained 1.0µg of probe (0.2µg/µl), 50% deionized formamide (Sigma), 10% sodium dextran sulphate (Sigma), 4.2µg ssDNA and 2XSSC, then mixed with excess blocking DNA of inbred line 330 which was 10-50µg respectively based on different tested ratios. Subsequently the hybridization mixture was denatured at 100 C for 10min, immediately chilled on ice for 10min. Hybridization was performed overnight at 37 C. The slides were then washed in formamide (20%), 2XSSC, 0.1XSSC, at 42 C for 15min each step, and subsequently washed in 0.1% TritonX-100, PBS at RT, for 5min each step.
DAB detection was performed according to the procedure outlined by Gustafson et al. (1990), Gustafson and Dille (1992).
The procedure included the following steps: 1) after the washes, the slide was covered with 30 µl (10 ng/µl) anti biotin FITC conjugate developed in goat (Sigma) and incubated in a humid chamber at 37 C for 45 min and the slides were washed 3 times, each for 5 min in PBS at RT; 2) subsequently covered with 30µl (11ng/µl) anti-goat-IgG conjugated with biotin (Sigma) instead, and incubated in a humid chamber at 37 C for 45 min and the slides were washed 3 times, each for 5 min in PBS at RT; 3)repeated step 1; 4) the slide was mounted in 20µl p-phenylenediamine dihydrochloride(10µg/ml) containing 3µg/ml propidium iodide (PI). Slides were analyzed using an Olympus fluorescence microscope.
An average of the hybridization site measurements was taken by calculating the distance from the centromere to the detection site and using that as a percentage of the arm on which the site was located. The arm ratio of the chromosome showing a detection site was also measured in order to determine on which chromosome the site was located. The standard deviations for both the percentage distance of the hybridization site and the arm ratio were calculated.
Different blocking ratios of the labeled DP genomic DNA: unlabeled maize Zi330 genomic DNA including 1:10, 1:20, 1:30, 1:40, and 1:50 were tested. Many scattered signal spots were distributed irregularly on different chromosomes and interphase nuclei of the tested pure line 540, Yi Dan 6 and the control, maize Zi330 with the lower blocking ratio including 1:10 and 1:20. The signals were not easy to detect even on chromosomes of the tested 540 and Yi Dan 6 with the higher blocking ratios 1:40 and 1:50. The optimum blocking ratio is 1:30. The following regular and stable results were observed through the eight repeated experiments with the blocking ratio of 1:30.
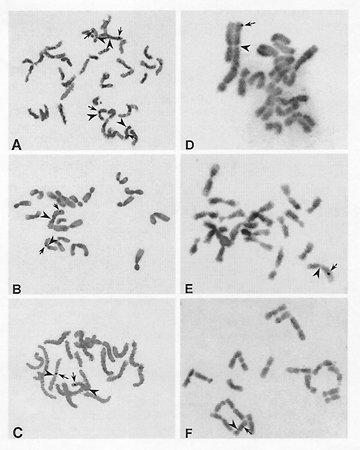
The genomic in situ hybridization signals of the pure line 540 appeared on the long arm of chromosomes 1 (Fig. 2A and C) and 8 (Fig. 2A), and chromosomes 2 (Fig. 2B) and 5 (Fig. 2A, 2B and C). Their detection rates were 10.10%, 11.15%, 8.01%, and 13.24% respectively and the percentage distances from centromeres to the hybridization sites were 86.3±4.5, 68.3±4.4, 12.2±2.5, 73.7±3.0 for the long arms of chromosomes 1, 2, 5, and 8 respectively (Table 1 and Fig. 2). In some mitotic cells, the signals showed simultaneously on the two members of the detected homologous chromosome pairs 1, 2, 5, and 8, each of the two members of chromosome 8 showing the signals at the same region can be visualized in Fig. 2A.
Table 1. Introgressed chromatin location
and arm ratio average of the chromosomes detected by GISH with the blocking
ratio of 1 probe: 30 Zi330 DNA
| Materials | arm |
|
|
Average percent distances of DAB and FISH detection (%) | Ratio of long to short arm | |||
| No. of detection | Detection rate (%) | Percent
distances (%) |
No. of detection | Percent
Distances (%) |
||||
| Alloplasmic pure line 540 | 1L** | 29 | 10.10 | 90.23±8.54* | 5 | 82.42±4.45 | 89.08±8.03 | 1.18±0.03* |
| 2L | 32 | 11.15 | 73.77±3.48 | 9 | 62.85±7.56 | 71.37±4.32 | 1.48±0.05 | |
| 5L | 23 | 8.01 | 14.25±1.75 | 6 | 10.23±3.32 | 13.42±2.04 | 1.04±0.02 | |
| 8L | 38 | 13.24 | 75.02±5.01 | 8 | 72.41±1.12 | 74.57±4.39 | 3.05±0.04 | |
| Yidan6 | 1L | 13 | 6.07 | 81.18±6.72 | 4 | 78.24±7.78 | 80.49±6.93 | 1.27±0.04 |
| 2L | 12 | 5.61 | 64.29±7.26 | 6 | 56.56±8.44 | 61.71±7.62 | 1.46±0.07 | |
| 8L | 15 | 7.01 | 48.85±5.52 | 6 | 41.61±3.49 | 46.78±4.99 | 2.98±0.02 | |
* Standard deviation, ** Long arm
The hybridization signals of Yi Dan 6 showed on the long arm of only one of two members for each of chromosome 1 (Fig. 2D), chromosome 2 (Fig. 2E), and chromosome 8 (Fig. 2F). No chromosome pair showed the signals simultaneously on their two homologues in all the mitotic cells observed. The detection rates of the signals were 6.07%, 5.61%, and 7.01%, and the percentage distances of the hybridization sites were 80.7±3.3, 60.4±2.9, and 45.2±4.4 for chromosomes 1, 2, and 8 respectively (Table 1). The control, maize, Zi330 did not show any hybridization signals on its chromosomes.
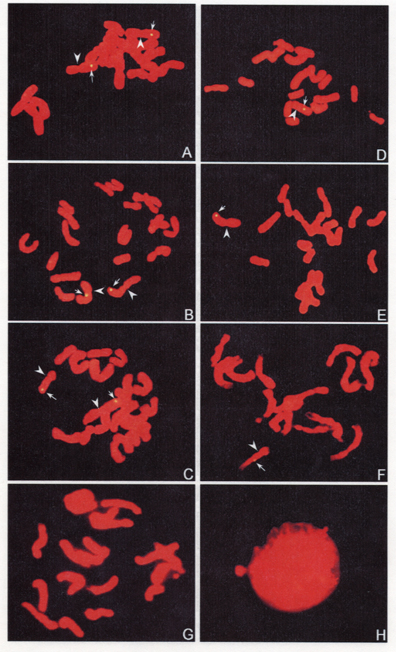
The signals observed by our fluorescence detection procedure showed yellow-green, while the chromosomes were red (Fig. 3). The blocking ratio was 1 labeled DP genomic DNA : 30 unlabeled maize Zi330 genomic DNA, just like that of DAB detection. The hybridization signals of the alloplasmic pure line 540 appeared on the long arm of chromosome 2(Fig. 3A), chromosomes 1 and 5 (Fig. 3B and C), chromosome 8 (Fig. 3C). In some mitotic cells, the signals showed simultaneously on the two homologues of the detected chromosomes 1, 2, 5, and 8. Each of the two members of chromosome 2 showing the signals at the same region can be seen in Fig. 3A. The percentage distances from centromeres to the hybridization sites were 82.42±4.45, 62.85±7.56, 10.23±3.32, and 72.41±1.12 for the long arms of chromosomes 1, 2, 5, and 8 correspondingly (Table 1).
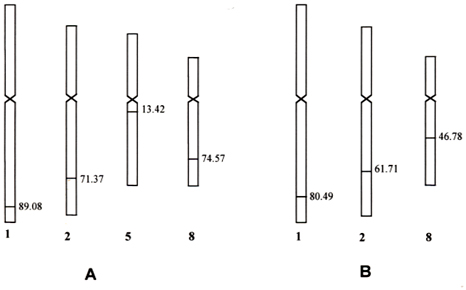
Compared with the results obtained by DAB and FISH detection systems, the chromosomes introgressed by DP chromatins were the same and the signal positions showed on the detected chromosomes, the percentage distances were very close to each other and had no significant differences statistically in both the pure line 540 and Yi Dan 6. Therefore, we have constructed the cytogenetic maps of the introgressed segments in both pure line 540 and Yi Dan 6 with the average value of the data obtained by two kinds of detection methods (Table 1 and Fig. 4).
The detection rates observed by FISH were about 30% for each hybridization site of the pure line 540, and about 20% for Yi Dan 6 with the blocking ratio 1:30. There are much higher than those by DAB detection.
The hybridization signals of Yi Dan 6 showed on the long arm of chromosome 2 (Fig. 3D), chromosome 1 (Fig. 3E), and chromosome 8 (Fig. 3F). The signals only appeared on one member of each detected chromosome pair and no signals on the two members of a chromosome pair were observed simultaneously in all the mitotic cells observed. The percentage distances of the hybridization sites were 78.24±7.78, 56.56±8.44 and 41.61±3.48 for the long arms of chromosomes 1, 2, and 8 respectively (Table 1).
No signals were observed on both the metaphase and interphase of the control, maize Zi330 (Fig. 3G and H).
In the pure line 540, the signals could be observed on two members of each detected chromosome pair, while in Yi Dan 6, only on one member. That the signals showed only on one member of each detected chromosome in Yi Dan 6 demonstrated that only one of its parents, 540, was introgressed, while the other, maize inbred line Zi 50003, had no introgressed DP chromatin at all. One member showing the signal must come from 540, the other from Zi50030 for chromosomes 1, 2, and 8.
It has been demonstrated that the stability of transgenes was related to the position integrated in transgenic plants (Jorgensen and Andersen 1994; Frello et al. 1995; Pedersen et al. 1997). Iglesias et al. (1997) reported that the transgenes integrated on the regions close to the telomeres were more stable. In 540, the alien chromatins of chromosomes 1, 2 and 8 were located in the regions close to the telomeres and their percentage distances from centromere to the hybridization site were over 70 (Table 1, Fig. 4), while those of chromosome 5 were located in the regions distal to the telomeres and the percentage distance was 13.42±2.07. In the genomes of both 540 and Yi Dan 6, the introgressed segments were located in the long arms on chromosomes 1, 2, and 8 except of chromosome 5 on which the signals only appeared in pure line 540 instead of Yi Dan 6. This meant that the DP chromatins integrated on chromosomes 1, 2, and 8 could be inherited stably, but the alien chromatins of chromosome 5 were unstable. Probably distant hybrids also follow the rule suggested by Iglesias et al. (1997) in transgenic plants. The alien chromatins integrated in regions distal to the telomeres on chromosome 5 might be lost during the processes of fertilization or hybrid seed production. Therefore, Yi Dan 6 had no signal on chromosome 5 at all.
In addition, the percentage distances of the signals showed smaller differences on chromosomes 1 (89.08 and 80.49) and 2 (71.37and 61.71), but they were more distinct on chromosome 8 (72.41 and 41.61) between 540 and Yi Dan 6 (Table 1). Because different regions of the chromosomes have varying states of condensation during the mitotic process, it is possible that the percentage distance of a giving hybridization site from the centromere will show some variation in different mitotic stages. For the differences of percentage distances on chromosomes 1 and 2 it could be explained by the fact that the observed samples could not be kept in the same mitotic stages between 540 and Yi Dan 6. Whether it was induced by chromosome rearrangement (translocation or inversion) or not in Yi Dan 6 for the larger position change on chromosome 8 remains to be studied.
In our laboratory, several important disease resistance genes of maize including Helminthosporium turcicum Pass, Helminthosporium maydis Nisik, and Helminthosporium carbonum Ullstrup resistance genes ht, rhm and hm with RFLP markers flanking each of them were mapped by in situ hybridization. The results showed that the hybridization site of ht1 was between the percentage distances 61.78 and 63.01 on the long arm of chromosome 2 (Li et al. 1998a); the hybridization site of homologous sequence of hm1 was located at 58.23 on the long arm of chromosome 5 (Li et al. 1998b); and that of the homologous sequences of rhm was positioned at percentage distances 65.06 and 72.40 on chromosomes 1 and 8 correspondingly (unpublished data). This demonstrated that all of the chromosome arms integrated by the alien chromatins had the distributions of the different disease resistance genes mentioned above in 540. Moreover, the sites of the disease resistance genes or their homologous sequences were very close to those of the alien chromatins on chromosomes 1, 2, and 8 except chromosome 5 on which the homologous sequence of hm1 was located at percentage distance 58.23, while the alien chromatin was at 13.42 (Table 1). As mentioned above, both 540 and the F1 hybrids, Yi Dan 6, showed much stronger Helminthosporium turcicum and Helninthosporium maydis resistances and wider adaptation to stressful conditions. The regions integrated by DP chromatins basically corresponded to those located by Helminthosporium turcicum and Helminthosporium maydis resistance genes ht and rhm on chromosomes 1, 2, and 8. Therefore, we think genes ht and rhm between maize and DP should be homologous or homoeologous, and the DP chromatins were integrated by homoeologous or homologous recombination from crossing over during meiosis instead of random process in the hybrids of maize X DP. Because the unexpressed introns should have different sequences in the genes between DP and maize, the introgressed DP chromatins could not be blocked easily with the blocking ratio 1 : 30. Because the differences of the introns between DP and maize were not absolute, the integrated DP chromatins were still blocked to a certain degree by maize genomic DNA with a high blocking ratio. Therefore, the signals of DP chromatins could not be detected completely with 1:40 and 1:50 blocking ratios. DP is interfertile with maize (Iltis et al. 1979), during meiosis the synapsis of their hybrids should be normal basically. Thus, many DP chromosome fragments or genes besides genes ht and rhm should be able to be integrated in the maize genome by crossing over. The alloplasmic pure line 540 was obtained by selection through several generations. As a result, it only showed the integration sites which were mainly corresponding to the loci of disease resistance genes in maize.
The breeding of alloplasmic pure lines, which showed disease resistance traits, offered us a new opportunity for establishing the specific gene library and isolating genes or chromatins specifying important agricultural characters, for example, resistance to disease and pests from wild germplasm by microdissection of the special regions on chromosomes. Based on our results with GISH, the establishment of the specific library containing genes ht and rhm of DP and their isolation could probably be performed by the microdissection of the DP integrated sites on the chromosomes of 540 or Yi Dan 6.
Acknowledgments: This research
was supported by the National Natural Science Foundation in China and the
Doctorate Vesting Point Foundation of the Education Ministry of the People�s
Republic of China
Return to the MNL 74 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}