In previous works we reported the isolation of lines of high and low B-chromosomes transmission rate (B-TR). This fact demonstrates the presence of genes controlling B-TR by both male and female sides (Rosato et al., Am. J. Bot. 83(9): 1107-1112, 1996). Recently, we made f.0B x m.2Bs crosses within and between high and low B-TR lines. We found that the resulting male B-TR depends on high or low B-TR line which the 0B female parent belongs to. This means that a high B-TR will be obtained if the female progenitor belongs to the high B-TR line (or a low B-TR if it belongs to a low B-TR group) independently of the origin of pollen (high or low B-TR lines). We concluded that B-TR is genetically controlled by the female parent without Bs, and these genes are located in the regular A chromosome set (Chiavarino et al., Am. J. Bot. 85(11): 1581-1585, 1998).
These results drove us to ask the following questions: Could B-chromosomes in the female progenitor affect the action of genes controlling the male B-TR? If so, do Bs enhance or repress the action of these genes? With the aim of evaluating these questions we crossed f.2Bs x m.2Bs and f.0B x m.2Bs (as control cross) only within high and low B-TR lines. The starting native population for selection was VAV 6313, race Pisingallo, with a high frequency of B-chromosomes (Rosato et al., Am. J. Bot. 85(2): 168-174, 1998). Lines used for this experiment suffered four generations of selection for high (0.82) and low (0.48) male B-TR.
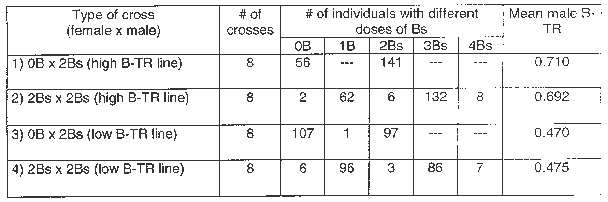
Eight successful crosses per each type of cross (a total of 32) were obtained and classified as: (1) f.0B x m.2Bs (both high B-TR line -control-), (2) f.2Bs x m.2Bs (high B-TR line), (3) f.0B x m.2Bs (low B-TR line, -control-), and (4) f.2Bs x m.2Bs (low B-TR line). The dose of Bs was determined in primary root tips of 19 to 30 individuals of each ear obtained to calculate B-TRs. The root tips were pretreated with 0.002 mol/L 8-hydroxyquinoline for 3 h at 20 C-22 C and subsequently fixed in ethanol:acetic acid (3:1). Finally, root tips were squashed in propionic hematoxylin (2%) with ferric citrate as a mordant.
We calculated the male B-TR in both types of crosses as the quotient between the mean number of Bs transmitted to the progeny by the male progenitor and the dose of Bs of the male progenitor (2Bs in all cases). The transmission of 2Bs in the female progenitor was mostly Mendelian, resulting in 1B in each egg cell. The male B-TR of the resulting f.2Bs x m.2Bs crosses was calculated considering only Bs transmitted by the male progenitor. Therefore if the progeny presents 1B and 3Bs, then the male progenitor transmitted 0B and 2Bs respectively and the female progenitor transmitted 1B in both cases. Few plants had 0B, 2Bs or 4Bs, their frequencies being 0.02, 0.02, and 0.04 respectively (Table 1). These data are explained by a failure in the formation of bivalents in the female progenitor. The result is two B-univalents that may migrate to the same pole. Plants with 0B may be formed since 2Bs migrate to the nonfunctional pole (nonfunctional megaspore) and the egg cell is fertilized by a sperm nucleus (SN) without Bs. Plants with 2Bs may be the result of migration of 2Bs to the functional pole (functional megaspore) and fertilization with a SN without Bs, or the migration of 2Bs to the nonfunctional pole and fertilization by a SN with 2Bs. In this case, it is impossible to determine whether Bs were transmitted by the male or female parent. Finally, plants with 4Bs may be formed if 2Bs migrate to the functional pole and are fertilized with a SN with 2Bs.
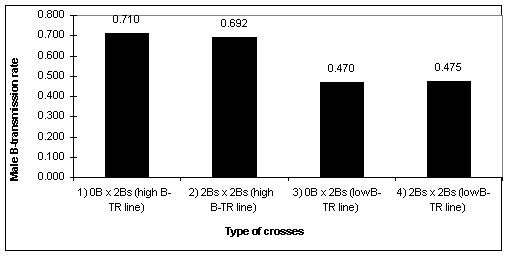
The high and low mean male B-TR were compared with a two-way ANOVA. There were significant differences between high B-TR and low B-TR lines (type of cross: 1 and 2 vs. 3 and 4; Figure 1) (F = 31.463, P < 0.00001), whereas no significant differences were found either between type of cross (F = 0.051, P = 0.8231), or interaction (F = 0.094, P = 0.7609) (Table 1, Figure 1). The results of this work indicate that in the low B-TR line, in fact, the B-TR is Mendelian (B-TR ª0.5), Bs being randomly transmitted to the progeny. High B-TR lines show a strong drive (B-TR > 0.5), irrespective of the presence of one B chromosome in the egg cell. In view of this, we dismiss the possibility that the presence of B-chromosomes in the egg cell affects the function of genes controlling male B-TR. Otherwise, Rosato et al. (1996) determined, by crosses f.1B x m.0B, that female B-TR is also genetically controlled on the female side. They also obtained two lines of high and low female B-TR. Considering these results, we concluded that there would be an equilibrium that keeps the frequency of individuals with Bs in the population. This hypothetical equilibrium would be reached by a balance among "anti-B" genes, causing a reduction in female B-TR, and "pro-B" genes, causing an increase in male B-TR, but controlled by the female progenitor genotype in both cases.
Table 1. Type of crosses with their mean B-transmission rate (B-TR).
Figure
1. Mean male B-transmission rate of the different types of crosses,
F=31.463, p<0.00001.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}