One of the main reasons for "incrossability" of different ploidy plants of one and the same species or close species is seed abortion conditioned by endosperm non-development. This phenomenon has been known for more than 70 years and for a long time hypotheses explaining it have been reduced to three possibilities:
1. deviation from the 2 : 3 : 2 ratio between numbers of maternal tissue endosperm and embryo genomes respectively (Muntzing, A, Hereditas 14:27- 42, 1930);
2. deviation from the 3 : 2 ratio of endosperm genomes to that of embryo (Watkin, AE, J. Genetics 25:125-162, 1932);
3. deviation from the 2 : 3 ratio of number of genomes in maternal cells to that of endosperm (Valentine, DM, Proc. 8 Intern. Botany Congress, Paris, p.170, 1954).
However as experimental methods of genetics progressed it was getting more and more obvious that the reasons for the anomalous development of endosperm are in its own genotypic structure (Johnston, SA et al., Theor. Appl. Genet. 57:5-11, 1980).
Classic experiments by Lin and the hypothesis formulated by him of parental imprinting of genomes in maize became the turning-point in our understanding of genetic mechanisms controlling the normal morphogenesis of endosperm (Lin, B-Y, Genetics 100:475-486, 1982; Genetics 107:103-115, 1984). It lies in the fact that values of expression of developmental genes introduced by maternal and paternal genomes are unequal. Therein any deviation from the ratio of two maternal genomes (2m) : one paternal (1p) leads to imbalance and seed abortion.
By now, thanks to Birchler and other researchers' works, we understand that besides imprinting, endosperm development is also influenced by other factors, but to all appearances imprinting is the main factor among them (Birchler, JA, Ann. Rev. Genet. 27:181-204, 1993).
Our interest in this phenomenon has to do with work on transfer of apomixis (pseudogamous type diplospory) from Tripsacum to maize where we met with a problem of seed size in apomictic intergeneric hybrids and with high sterility partly associated with ovary death in early stages of development. In a work published by Daniel Grimanelli with co-workers (Sex. Plant Reprod. 10:279-282, 1997) absence of imprinting effect in crossing diploid and tetraploid Tripsacum species (Poaceae) was shown. The same was also evidenced by the results obtained at our laboratory earlier when no dosage effects were observed in apomictic BIII-hybrids of maize with Tripsacum with 2 to 9 maize genomes accumulated (Petrov, DF et al., Apomixis and its role in evolution and breeding, New Delhi, India, 9-73, 1984; Kindiger, B and Sokolov, VA, Trends in Agronomy, 1:75-94, 1997).
The material for this research was progeny of an F1 hybrid obtained in 1964 from crossing tetraploid maize with Tripsacum having a genome formula 2n=56 (20Mz + 36Tr). As a result of backcrossing, numerous BII and BIII-hybrids with a different ratio between complete parental genomes or between complete maize genomes and Tripsacum subgenomes or separate chromosomes were derived. In this case the term "subgenome" was given by us to a stable group of 9 chromosomes of the wild parent (obtained in one of the BII-hybrids) the carriers of which have been steadily exhibiting for 20 years the same apomixis as the hybrids with a complete genome of 18 chromosomes. The pedigrees of these hybrids were published earlier (Sokolov, VA et al., Russian Genetics 34:499-506, 1998).
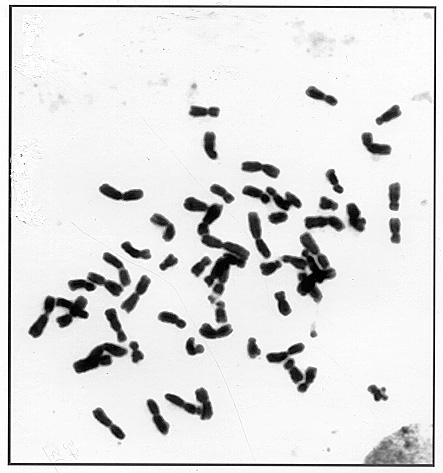
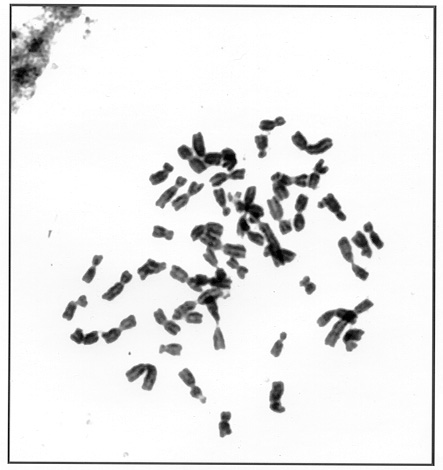
The exception is a line with 63 chromosomes developed from backcrossing a 29-chromosome BII-hybrid (20Mz + 9Tr) in one of its generations. This BII-hybrid pedigree will be reported by us in a separate publication later. Here we'll only mention that the 63-chromosome twins originated from a 43-chromosome plant as a result of fertilization by tetraploid pollinator of unreduced egg cell (BIII-hybrid) (see Fig. 1). From these twin plants after pollination with tetraploid some 50 filled but small size seeds - subnormal by Lin's terminology - were obtained by us. Unfortunately only part of them were viable and all plants obtained from them were BII-hybrids with 50 maize chromosomes and an addition of 3 (about 50% - Fig. 2) to 1 or 2 Tripsacum chromosomes. They were not totally female fertile and so only part of them produced seeds, and only when pollinated with tetraploid. The cytogenetic structure of this generation has not been analysed yet.
Figure 1. The BIII-hybrid 2n=63 (60Mz + 3Tr).
Figure 2. The BII-hybrid from 2n=63, 2n=53 (50Mz + 3Tr).
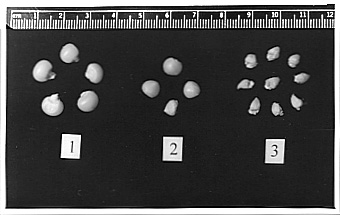
Apomixis in gamagrass and, therefore, in the hybrids is pseudogamous diplospory of the Antennaria-type. So for the kernel to develop normally fertilization of the polar nucleus is needed. The results of pollination with tetraploids (pollinator 2n=4x) and diploids (pollinator 2n=2x) are reflected in the ratios between maternal and paternal genomes presented in the table. The relative seed size, we put down in the column "normal" of the table, in Fig. 3 is designated by numbers 1 and 2, the "subnormal" size is indicated by number 3.
Figure 3. The relative seed size: 1, 2n=39 (30Mz + 9Tr); 2, the offspring of 2n=51 (50Mz + 1Tr); 3, 2n=53 (50Mz + 3Tr).
Table.
| Genotypes of apomictic line | Ratio between genomes in endosperm pollinated with | Seed size | |||
| tetraploid | diploid | Normal | Subnormal | ||
| 38 20Mz + 18Tr | 4m : 2p | 4m : 1p | + | ||
| 48 30Mz + 18Tr | 6m : 2p | 6m : 1p | + | ||
| 58 40Mz + 18Tr | 8m : 2p | 8m : 1p | + | ||
| 39 30Mz + 9Tr | 6m : 2p | 6m : 1p | + | ||
| 49 40Mz + 9Tr | 8m : 2p | 8m : 1p | + | ||
| 59 50Mz + 9Tr | 10m : 2p | ? | + | ||
| 79 70Mz + 9Tr | 14m : 2p | ? | + | ||
| 78 60Mz + 2 x 9Tr | 12m : 2p | ? | + | ||
| 96 60Mz + 2 x 18Tr* | 12m : 2p | - | - | - | |
| 63 60Mz + 3Tr** | 8m : 2p | ? | + | ||
| 53 50Mz + 3Tr | 6m : 2p | ? | + | ||
*A dwarf plant does not eject silk, does not set seeds
**Nonreduction without parthenogenesis
As follows from the results presented in the table the varying of the ratio between maize genomes of maternal and paternal origin within a wide range after pollination with tetraploid (from 4m : 2p to 14m : 2p) as well as diploid (from 4m : 1p to 8m : 1p) does not lead to imprinting manifestation, and both endosperm and embryo develop normally producing viable seeds. In cases when the lines were not pollinated with diploid, and so we lacked appropriate results from this manipulation, this fact is marked as "?" in the table. There's no doubt that this is observed by reason of the presence in the hybrids of a complete Tripsacum genome or its part of 9 chromosomes in which case "normal" endosperm develops. With three Tripsacum chromosomes available in the BIII-hybrid (2n=63) and then in its reduced progeny (the BII-hybrid 2n=53), abnormally small "subnormal" endosperm is formed.
With a very low frequency plants with genome duplications (2n=78; 2 x 30Mz + 2 x 9Tr and 2n=96; 2 x 30Mz + 2 x 18Tr) are encountered. They appear to be a result of lack of one single megaspore division in rare cases of meiotic diplospory in Tripsacum. The plant having 96 chromosomes developed abnormally - it was dwarf, with a very little ear, did not eject silk and produced no progeny. From the 78-chromosome plant not differing in phenotype from the 39-chromosome sibs, with its sufficiently high sterility, we managed to obtain seeds and these were subnormal in size.
Thus, based on the results presented, one may conclude that the availability of Tripsacum chromosomes in some way inhibits imprinting expression in the hybrids. And it's quite obvious that for its total suppression some critical number of Tripsacum chromosomes is needed. In this stage of the research we can assert the validity of this conclusion for 9 chromosomes.
The development of the subnormal size grains obtained from the 78-chromosome plants may be explained by imbalance between maize genomes and twice the set of 9 Tripsacum chromosomes. It need be noted that despite twice the chromosomes these plants are male sterile, moreover the development of their anthers is disturbed and meiosis does not occur in the anthers.
The research was supported by Russian Foundation of Basic Investigation
Grant No. 97 - 04 - 49301.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}