In a previous publication it was shown that crossing two haploid-inducing lines, KMS and ZMS, resulted in the production of transgressive genotypes whose haploid-inducing capacity was more then two times higher than that of the parental lines (Chalyk et al., MNL 68:47, 1994). It was suggested that the parental lines KMS and ZMS differed from each other by two genes controlling haploid-inducing capacity. Transgressive genotypes carry both genes and are capable of inducing 7 to 9% of haploids.
The objective of our subsequent work was to produce new homozygous haploid-inducing lines which would be superior to the parental lines KMS and ZMS in haploid-inducing ability and would carry marker genes A1; B1; C1; R-nj and Pl1 allowing haploids to be identified both at the stage of mature seeds and at the stage of vegetating plants.
The parental line ZMS carries marker genes A1; C1; R-nj, which enable haploids to be identified at the stage of mature seeds by the anthocyanin coloration of the top of the endosperm and lack of coloration in the embryo. The parental line KMS carries genes a1; B1; Pl1 which allow haploids to be identified at the stage of 3 to 5-day-old seedlings by the lack of anthocyanin coloration in roots (Tyrnow and Zavalishina, DAN 276:735-738, 1984). Both of the haploid inducers, ZMS and KMS, were created on the base of Coe�s line, Stock 6 (Coe 1959).
We worked with the progeny resulting from selfing of the KMS x ZMS cross. Selfed in each generation were only those plants which exhibited intense anthocyanin coloration. Upon harvesting, the ears were examined for the presence of R-nj. Up to generation F8, individual plants were evaluated for two major traits: (1) haploid-inducing capacity, and (2) intensity of the embryo, endosperm and whole plant coloration. In F2 and F3, the haploid inducing capacity was measured by the number of haploids induced within the lines. Starting with F4, the haploid inducing capacity was evaluated by applying pollen from the selfed plants to diverse genetic material: lines, hybrids, synthetic populations.
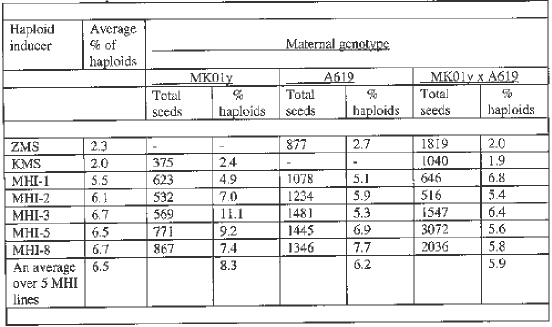
In 1998, fourteen lines of the F8 progeny were sown in the field. Their perfect uniformity suggested that all of them were highly homozygous. Subsequent evaluation was, therefore, performed for a line as a whole rather than individual plants. For estimating the haploid-inducing capacity, a mixture of pollen from several plants of the evaluated inducer line was taken. Used as a maternal parent were two inbred lines, MK01y and A619, and a cross between them, MK01y x A619. The results obtained for the best five new haploid inducers and parental lines ZMS and KMS are presented in Table 1.
New haploid inducers, MHI (Moldovian Haploid Inducer), were superior to the parental lines ZMS and KMS. Five MHI lines induced an average of 6.5% haploids over three maternal genotypes. The percentages of haploids induced by them were approximately equal, ranging from 5.5% to 6.7%. This was almost three times higher than the frequency of haploids induced by initial haploid-inducing lines, ZMS and KMS. Weak expression of marker genes in MK01y x ZMS and A619 x KMS did not allow us to select haploids from this material, while we had no problem with expression of the marker system when our new haploid-inducing lines were applied. The new inducers did not differ from one another phenotypically: the plants were dark violet; the R-nj gene was clearly expressed in the seeds.
Table 1. Frequency of seeds with haploid embryo resulting from pollination with pollen from maternal haploid inducers.
The maternal genotype had a valuable effect on the frequency of haploids produced. The highest percentage of haploids was obtained with line MK01y - an average of 8.3%. In line A619, an average output of haploids was 6.2%. The MK01y x A619 hybrid yielded the lowest percentage of haploids - 5.9%.
We suggest that one of the factors determining the proportion of haploids of a particular maternal genotype is the coinciding or differential periods of flowering of the maternal genotype and the haploid-inducing line. In our experiment, the MK01y x A619 hybrid started silking 5 days before pollen shedding in haploid-inducing lines, whereas in line MK01y the silking coincided with mass pollen shedding in haploid inducers. The line A619 started silking 2-3 days before pollen shedding in haploid-inducing lines.
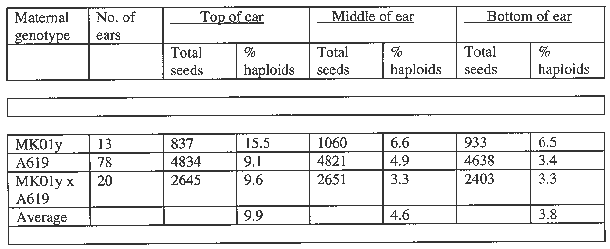
Previously, it has been established that the frequency of occurrence of haploids is not the same in different portions of the ear (Chase, Amer. J. Botany 35:117-167, 1969). The highest proportion of haploids occurred in the top portion of the ear where the egg cells are the youngest. We also measured the frequency of haploids in three ear parts: top, middle and bottom. The results are presented in Table 2.
Table 2. The frequency of seeds with haploid embryo occurring in different portions of the ear.
Our results strongly support the conclusions reached by S.Chase about thirty years ago. The young egg cells in the top portion of the ear are more capable of developing without fertilization and of giving rise to a haploid embryo than the older egg cells in the bottom of the ear.
Involved in the above analysis were all the ears of line A619 resulting from crosses with haploid inducers and some ears of line MK01y and the MK01y x A619 hybrid. As averaged over three maternal genotypes, the frequency of haploids in the top portion of the ear was 9.9%. This is almost three times the frequency of haploids in the bottom of the ear and more than twice that in the middle portion. The results in Table 2 and those obtained by S.Chase allow the supposition that the proper timing of pollination of ears may increase the output of haploids.
From the results obtained in our experiment it may be concluded that
new inducers of maternal haploids have been created which allow haploids
to be produced in large numbers from almost any genotype of maize.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}