Genetic data suggest that Ac transposes in maize during replication. Based on the observation that Ac transposase (TPase) binds in vitro selectively to hemimethylated DNA and that the TPase binding sites of Ac/Ds are methylated in vivo, we have proposed a model that explains the association of transposition with replication, and the chromatid selectivity of transposition as a consequence of different Ac/Ds methylation states (Wang, L et al., Plant Cell 8:747-758, 1996).
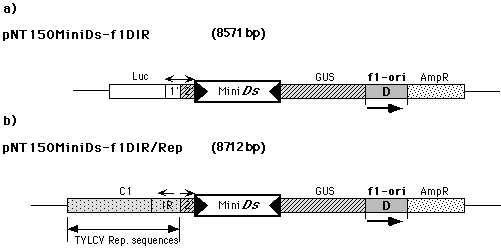
To validate this model, we have begun to investigate the effect of replication on Ac/Ds transposition in the petunia protoplast test system. We constructed two reporter plasmids. In pNT150MiniDs-f1DIR (Fig. 1a) a 700bp long Mini-Ds element blocks the expression of the GUS gene. Plasmid pNT150MiniDs-f1DIR/Rep (Fig. 1b) is identical to pNT150MiniDs-f1DIR, except that the 1�-Pr./LUC gene fragment has been replaced by a viral replicon from the Tomato Yellow Leaf Curl Virus (TYLCV) (Kheyr-Pour, A et al., Nucleic Acids Res. 19:6763-6769, 1992).
To determine the transposition activity, each reporter plasmid was co-transfected together with a TPase expression plasmid into ca. 106 petunia protoplasts. The TPase protein enables the Ds element to transpose, resulting in reversion of the GUS gene. After transfection, protoplasts are cultured for approximately 30 hr in 5 ml rich medium. Subsequently, 200 µl of the protoplasts are spread and fixed on nitrocellulose membranes, and stained for GUS activity. The number of blue protoplasts is taken as an approximation of the Ds excision frequency.
According to our preliminary results, in the presence of the TYLCV replicon the number of blue protoplasts is 2 to 3 times higher than without it (Table 1). Moreover, the staining intensity of the protoplasts is significantly higher after transformation with the replicon-containing plasmid. To determine whether this effect is merely the result of the altered sequence composition of the pNT150MiniDs-f1DIR/Rep plasmid, or if the replication activity of the TYLCV replicon is responsible for the apparent higher transposition frequency, we performed several tests.
(1) Petunia protoplasts were transfected with reporter plasmids isolated from Dam+-Dcm+ host strains. Control experiments using plasmids from dam-dcm- strains demonstrated that Dam-Dcm methylation does not affect the transposition efficiency of the Ds element in Petunia protoplasts. We determined the GATC methylation state of pNT150MiniDs-f1DIR and pNT150MiniDs-f1DIR/Rep isolated from a Dam+-Dcm+ strain after transfection into protoplasts. The restriction enzyme MboI exclusively digests DNA containing unmethylated adenine residues within its recognition sequence, GATC. Therefore, plasmid DNA is initially resistant to MboI digestion, whereas after a few replication cycles in Petunia cells, the plasmid becomes sensitive to MboI digestion. Total DNA was isolated from protoplasts 2, 4 and 6 days after transfection and was digested with MboI and a second, methylation-insensitive restriction enzyme that linearizes both plasmids. Southern blot hybridization analysis using a 1024 bp MboI fragment of the two plasmids as a probe showed that MboI digestion released the 1024 bp fragment in increasing amounts at 2, 4, and 6 days after transfection in pNT150MiniDs-f1DIR/Rep transfected protoplasts. However, this fragment was not detected in pNT150MiniDs-f1DIR transfected protoplasts. This result shows that only the plasmid containing the viral replicon becomes MboI-sensitive in transfected protoplasts, indicating that pNT150MiniDs-f1DIR/Rep actively replicates in the cells, but not pNT150MiniDs-f1DIR. These results suggest that DNA replication improves transposition of the Ds element, independently from the methylation status of its DNA.
(2) Our model predicts that fully methylated Ds should be unable to transpose in the absence of replication. After replication, supposedly only one of the two hemimethylated daughter elements is transposition competent. To test this model, we have treated pNT150MiniDs-f1DIR and pNT150MiniDs-f1DIR/Rep plasmids with SssI methylase that methylates all cytosine residues to 5mC within the double-stranded dinucleotide recognition sequence 5'...CG...3'. After the reaction all CpG dinucleotides in the plasmids, including the Ds element, are methylated. We have obtained preliminary results after transfection of protoplasts with C-methylated and unmethylated reporter plasmids and subsequent GUS staining (Table 1):
Table 1. Number of GUS positive protoplasts after transfection with
C-methylated and unmethylated reporter plasmids.
| Plasmid | No. of blue protoplasts |
| pNT150MiniDs-f1DIR | 170 |
| pNT150MiniDs-f1DIR/Rep | 450 |
| pNT150MiniDs-f1DIR-Met | 2 |
| pNT150MiniDs-f1DIR/Rep-Met | 230 |
The frequency of GUS-positive protoplasts with the non-replicating, fully C-methylated plasmid is almost zero, whereas the replicating plasmid gives rise to normal frequencies. The lack of GUS-positive protoplasts with pNT150MiniDs-f1DIR-Met can be explained by one of two mechanisms. Either, in accordance with our model the fully methylated Ds is unable to transpose in the absence of replication and thus the GUS gene remains blocked, or alternatively, the Ds can excise, but due to the methylation the 2�-promoter persists in an inactive state. Experiments are under way to distinguish between these mechanisms. Preliminary (yet unconfirmed) PCR data indicate that the Ds excision frequency is significantly reduced (more than one order of magnitude) in pNT150MiniDs-f1DIR-Met compared to pNT150MiniDs-f1DIR/Rep-Met.
We have begun to investigate the effects of hemimethylation on Ds transposition. Two pNT150MiniDs-f1DIR/Rep DNAs have been produced in which either one or the other DNA strand in the Ds element are CpG methylated, whereas outside Ds the plasmid is unmethylated. In the very first transfection experiment with these two DNAs, the Ds excision frequencies differed significantly. If these data can be confirmed, they strongly support our model.
Figure
1. a) pNT150MiniDs-f1DIR construct. b) pNT150MiniDs-f1DIR/REP
construct. Luc: Luciferase gene, f1-ori: origin of replication
of M13 bacteriophage, with D is just indicated the orientation of
the f1 replication origin, 2': GUS gene promoter, 1':
Luciferase gene promoter, MiniDs: 700-bp Ds element, GUS:
GUS gene, AmpR: ampicillin resistance, IR: region containing
the promoter and the DNA binding sites for the viral C1 protein, C1:
viral protein responsible for the replication function.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}