Previously, Greyson et al. (Devel. Genet. 18:244-253, 1996) and Yang et al. (MNL 72:56. 1998.) have demonstrated through in situ hybridization that the heat shock response in maize seedlings is localized to meristematic regions of primary roots, lateral roots and shoots. These findings raise questions about the nature of this localized response. To address this question we employed the use of in vitro callus cultures (Friedberg et al., MNL 72:56. 1998) to determine if the heat shock response requires an organized meristem or if actively cycling cells can respond. Furthermore, we characterized the heat shock response in not just the callus but in roots that were induced to regenerate from callus.
To assess the heat shock response in maize callus we employed the use of the antisense in situ hybridization, using the scMHSP 18-9-2 RNA, DIG labeled probe (Table 1, Greyson et al., 1996). All callus and root tissues were heat shocked at 43 C for two hours, fixed and embedded in wax blocks. Ten and 15 micron sections were prepared and mounted on glass cover slips in preparation for in situ hybridizations. All in situ hybridizations were performed according to Greyson et al., 1996. Callus was initiated from immature embryos (inbred Oh 43) and subjected to in situ hybridizations after 30, 60, 90 or 120 day intervals. At each of these intervals, several additional calli were transplanted to root induction media. Once root induction was evident (approximately 5-10mm), the roots were also subjected to in situ hybridizations with the aforementioned probe.
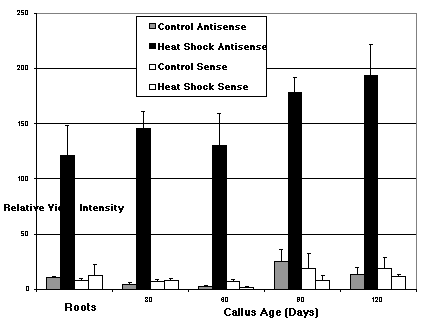
Analysis of callus in situ hybridizations revealed a significant level of HSP 18 mRNA expression in callus 30, 60, 90 or 120 days old in response to heat shock (Figure 2). The experiment was replicated eight times and there appeared to be no significant difference in HSP 18 mRNA expression between the intervals tested (Figure 1). Analysis of in situ hybridizations on the callus-induced roots exhibited positive HSP 18 mRNA expression, which was localized to cells in the meristematic region. Observations of heat shocked callus sections revealed that mRNA expression was localized to the nucleus and the periphery of the cells (Figure 2, E to H) and not throughout the entire cell as is seen in meristematic tissue (Yang, 1997, MSc Thesis The University of Western Ontario, London, Ontario). Although it could not be observed directly, the mRNA localization in callus might be a result of cells with large vacuoles, possibly restricting the mRNA to the periphery and the nuclear region. Furthermore, it is possible that some of the cytoplasmic mRNA could have been lost due to the rupturing of cells during sectioning.
The roots that regenerated from callus cultures appeared to possess the cell anatomy of embryo-derived roots and when heat shocked, a characteristic heat shock response. As the roots elongated and the cells differentiated the heat shock response diminished in the mature cells but was present in the meristematic regions. It appeared that the regeneration process did not affect the functioning of organized tissue.
It was observed that across the four time intervals the heat shock response remained at a constant level indicating that the cells were still alive and active. This provides further evidence that cells in callus culture have the ability to remain alive beyond the life span of an embryo-derived plant and may have the ability to remain active indefinitely.
The callus appeared to expand outward from the edges and not from the center. Analysis of callus sections revealed that cells throughout the callus have the ability to respond to heat shock. This may indicate that the cells inside the callus are actually dividing and the callus increased from both the outside and the inside. Alternative possibilities are that the cells were not dividing but retain the ability to express the heat shock response.
The probe used in these hybridization experiments was specific to the ORF of HSP 18 as confirmed by Northern blot hybridization. However, in maize there are three different HSP 18 genes (cMHSP 18-1, cMHSP 18-3 and cMHSP 18-9) that code for different HSP 18 mRNAs that share considerable homology in their ORFs (Goping et al., 1991, Plant Molec. Bio. 16:699-711; Atkinson et al., 1993, Dev. Genet. 14:15-26). Further, it is known now that there are multiple translation start sites yielding up to eight distinct proteins from these three genes (Frappier et al., 1998, Genet. 148:471-477). Thus, it is not known which HSP 18 mRNAs are expressed in heat shocked callus. Further studies using probes specific for the 3� and 5� untranslated regions should be employed to determine which of the HSP 18 mRNAs are expressed in heat shocked callus.
The evidence that callus can express a heat shock response indicates that the ability to respond to heat shock is not only a characteristic of meristematic tissue but a characteristic of any cells in an active nuclear cycle. Further support can be seen in the ability of secondary roots to express a strong heat shock response even though they emerge from mature cells that cannot respond to heat shock (Yang, 1997).
Figure 1. Densitometric readings of DIG stained in situ hybridization sections using the scMHSP 18-9-2 antisense and sense probes. A) Unadjusted data averaged from the relative video intensity of callus sections (30, 60, 90 and 120 days old) and root (induced from callus) sections. The data were drawn from control and heat shock sections hybridized with the antisense probe, and control and heat shock slides hybridized with sense probe. B) The average relative video intensity representing the net antisense heat shock response (control data subtracted from heat shock data) for each time interval and roots (induced from callus). There is not a significant difference among the time intervals of the callus response nor is the callus response significantly different from that measured in the roots. Error bars = the standard deviation; sample size (n) = 5.
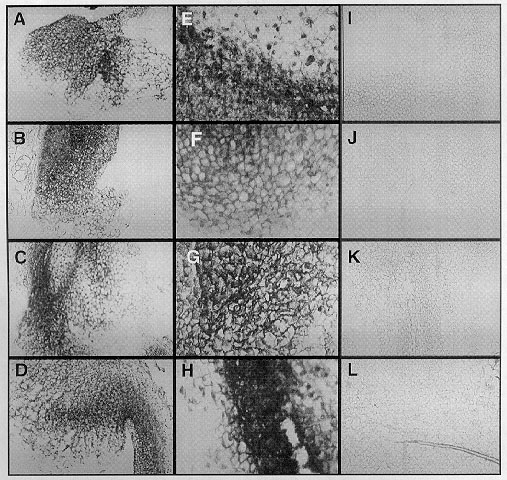
Figure
2. Reproduction of digital photographs of representative in
situ hybridizations of 30, 60, 90 and 120 day old heat shock and control
callus sections probed with the scMHSP 18-9-2 antisense RNA probe. A&E)
30 day old callus heat shocked at 43 C for two hours. B&F) 60 day old
callus heat shocked at 43 C for two hours. C&G) 90 day old callus heat
shocked at 43 C for two hours. D&H) 120 day old callus heat shocked
at 43 C for two hours. I to L are control callus from 30, 60, 90 and 120
day old callus respectively. Notice the increase of HSP 18 mRNA (dark regions)
in the heat shock callus (A to H) relative to the controls (I to L). Pictures
A to D and I to L were taken at 100x magnification. Pictures E to H were
taken at 400x magnification.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}