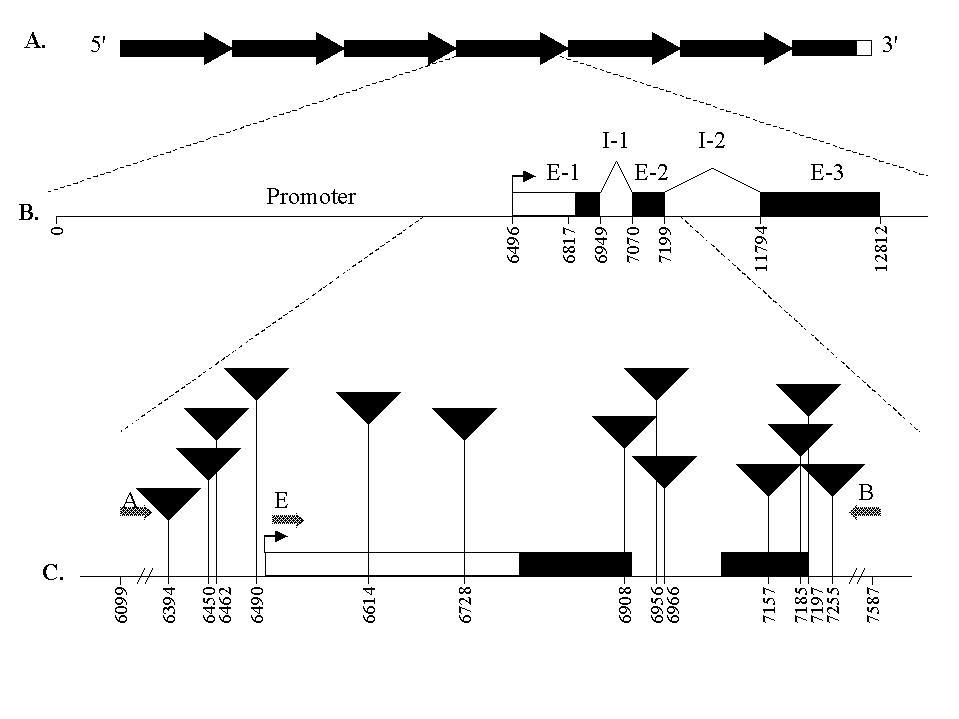
The maize P locus encodes a Myb-homologous regulator of red phlobaphene pigment biosynthesis in pericarp, cob glumes, and other floral tissues (Grotewold et al., Cell 76:543-553, 1994). The P-wr allele of the P gene specifies the accumulation of pigment in the cob, but not the pericarp (Styles and Ceska, Maydica 34:227-237, 1989). P-wr consists of a multi-copy gene complex with approximately six tandem repeats aligned in a head-to-tail arrangement (Figure 1A) (Chopra et al., Molecular and General Genetics 260:372-380, 1998).
In order to gain a greater understanding of the tissue specificity associated with this allele, induced mutations were identified using the Trait Utility System for Corn (TUSC) developed by Pioneer Hi-Bred International, Inc. (Meeley and Briggs, MNL 69:67,82; Bensen et al., Plant Cell 7:75-84, 1995). TUSC is a reverse genetics tool based on PCR and the Mutator transposable element family. Using several P-wr specific primers, we have detected 13 unique Mu insertion events within a 1.4 kilobase region of P-wr including a portion of the promoter, exon 1, intron 1, exon 2, and part of intron 2 (Figure 1C). Screening for insertion events in the 3' region of P-wr is continuing.
We have not yet identified which of the six copies of P-wr contain the Mu element insertions. There are a number of sequence polymorphisms in the P-wr genomic sequences, both among the segments flanking the 13 different Mu insertion clones, and in comparison to the previously determined genomic and cDNA sequences of P-wr from inbred line W23 (Chopra et al., Plant Cell 8:1149-1158, 1996). These polymorphisms include a twenty-eight base pair direct duplication, several smaller (3 � 6 bp) direct duplications, and numerous single base pair changes, insertions, and deletions. Many of the polymorphisms are found on more than one independent clone, ruling out their origin from PCR or sequencing errors. These polymorphisms could be due to differences between the several P-wr alleles that were used in the TUSC population, or to sequence differences among the six gene copies of the P-wr complex. The latter situation would facilitate the mapping of Mu insertions to a particular gene copy.
An unexpected outcome of the TUSC approach is the possible detection of the 3' end of the P-wr gene complex. The junction between the P-wr complex and the flanking chromosomal DNA has eluded previous conventional cloning attempts. One of the TUSC clones contains a Mu insertion located 157 bp 3' of a truncated P-wr sequence. This truncated P-wr copy contains the promoter region, but none of the coding sequence. The 157 base pairs between the truncation of P-wr and the Mu element have no homology to the known P-wr sequence, nor any relevant database hits. PCR experiments on genomic DNA have confirmed the presence of this copy, but a positive Southern blot confirmation has yet to be completed.
The phenotypes of the Mu insertion alleles should help to identify which copy or copies of the P-wr complex are functional. Gain of P function in pericarp and/or loss of function in cob glumes are particularly striking phenotypes that may result from transposable element insertions into the P-wr gene. The genetic transmission and/or phenotypes resulting from many of these Mutator insertion events are yet to be characterized, and one season of outcrosses may be necessary to make the plants more vigorous. However, one interesting case was identified in the summer of 1998. A plant with a gain in P pericarp function has been identified as an ear with a red sector. The ear was derived from a family that contains a Mu insertion in the 5' untranslated leader sequence (position 6728). The accumulation of pigment in the pericarp may be a result of the phenomenon of Mu suppression, wherein transcription begins from an outward-reading promoter present in the TIR of the Mu element (Barkan and Martienssen, Proc. Natl. Acad. Sci. USA 88:3502-3506, 1991). Further testing will be required to establish the mechanism of this ectopic P-wr expression in pericarp. Nonetheless, this result indicates that the P-wr gene product can function in pericarp when it is expressed there. Analysis of this and other novel phenotypes arising from the mutant plants should provide key insights into the tissue specificity associated with P-wr.
Figure
1. A. The P-wr multi-copy gene complex. Each 12.6 kb repeat
is indicated by the arrows. The 6.4 kb truncated copy lying at the 3' end
is represented by the dark box. The 157 base pairs of chromosomal DNA flanking
one Mu insertion is indicated as the putative 3' end of the P-wr
complex (open box). B. Enlargement of a single P-wr copy showing
the exon/intron structure. The dark arrow shows the transcription start
site, the open box indicates the untranslated leader sequence, and the
black boxes indicate the coding regions. C. Enlargement of
the area screened for Mutator insertions using TUSC. The triangles
represent Mu element insertion locations, with the nucleotide position
indicated below. The positions of PCR primers A, B, and E used for TUSC
pool screening are shown by the large arrows.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}