Previous work in this laboratory focussed on localizing B chromosome sites that control nondisjunction. Selection of mutants was helped by a screening technique with the translocation, TB-9Sb. In this procedure, a high nondisjunction genetic background was utilized and the relatively few kernels that showed a lack of nondisjunction for the B-9 were selected as potential mutants of nondisjunction (Carlson, Chromosoma 42: 127, 1973). A reasonable percent of kernels that lacked nondisjunction in the initial screen were found to be "mutants". The mutants turned out to be spontaneous deletions of different regions of the B chromosome, either on the B-9 or the 9-B. This method (in combination with the work of several other laboratories utilizing different techniques) helped localize sites on the B chromosome that were required for nondisjunction.
Selecting in the same way for mutants of preferential fertilization is not feasible. The problem is that there is no way to enrich the population studied for mutants. Instead, every plant in a population must be testcrossed to detect a lack of preferential fertilization. The number of crosses required is prohibitive. An alternate approach is to analyze existing deletions, found in the nondisjunction screen, for a lack of preferential fertilization. It seems likely that some of the deletions should overlap with sites controlling preferential fertilization. The fact that these deletions do not permit nondisjunction presents a problem, since nondisjunction is necessary for a measurement of preferential fertilization. A systematic study of these deletions, therefore, seems impossible. However, in one case the problem can be sidestepped.
A derivative of the B-9 chromosome of TB-9Sb was isolated in 1970 (Carlson, Chromosoma 30: 356). This derivative was shown to be a pseudoisochromosome, which presumably lacks the B short arm (if such an arm exists). The two arms of the pseudoisochromosome differ in the proximal region. One arm lacks the B centric heterochromatin and is incomplete. The other arm appears to be complete (Carlson and Chou, Genetics 97: 379, 1981).
The pseudoisochromosome (also called the original iso) misdivides frequently to give rise to telocentrics. The telocentrics lacking centric heterochromatin are called type 1 telocentrics, while the ones which have centric heterochromatin are called type 2.
Isochromosomes can also be recovered from the telocentrics by misdivision. For the purposes of this report, there are only a few types of chromosomes: the original iso, type 1 telos and isos and type 2 telos and isos. Studies by Kaszas and Birchler (Embo J. 15: 5246, 1996) use a different nomenclature for these chromosomes to indicate the number of misdivision events that each chromosome has undergone.
The type 2 telos are capable of high rates of nondisjunction and can be tested, without difficulty, for preferential fertilization. They show an excess of nondisjunctional kernels in which the B-9 telos are present in the embryo. In other words, they show preferential fertilization. For example, five ears were classified in a cross of yg/+ bz bz x 9-B 9-B type 2 telo B-9. The male parent is hemizygous for the B-9. All its viable meiotic products are: 9-B B-9. Nondisjunction is seen as bz kernels that give Yg seedlings and Bz kernels that give yg seedlings. The five ears in this cross produced 863 Yg bz kernels and 206 yg Bz kernels. The predicted ratio of Yg bz to yg Bz is 2:1 for random fertilization, rather than 1:1, since the tester is heterozygous for yg/+ and cuts the frequency of yg Bz kernels in half. The observed ratio was 4.2:1.0. This ratio would be even greater if the rate of germination were taken into account, since the bz class germinated at a rate of 89.5% while the Bz class germinated at a rate of 97.8%. (This variation is due to linkage of the detrimental sh phenotype with bz). The estimated rate of preferential fertilization in the cross, taking germination rates into account, is 69.6%. This is fairly typical for standard TB-9Sb crosses (Carlson, Genetics 62: 543, 1969) and suggests no impairment in functioning of the preferential fertilization system.
The type 1 telos are not so easily tested for preferential fertilization. They are incapable of nondisjunction, due to deletion of the centric heterochromatin. Therefore, the Yg bz and yg Bz classes do not ordinarily exist in crosses with the telocentric. However, this barrier to testing preferential fertilization can be overcome. It was shown recently that the type 1 telocentric is unstable when present in the hemizygous condition. Crosses of a yg yg bz bz tester as female to plants with 9-B 9-B type 1 telo B-9 produced significant numbers of Yg bz and yg Bz phenotypes. These resulted from misdivision of the centromere rather than nondisjunction (Carlson, MNL 70: 28, 1996). The hemizygote, therefore, can be used in a test of preferential fertilization.
Instability of the type 1 telocentric apparently arises in meiosis, through misdivision of the unpaired chromosome. Misdivision produces an unstable telocentric. If the unstable telocentric is transmitted to the second pollen mitosis, it can undergo a pseudo-nondisjunctional process. The telocentric can misdivide and produce an isochromosome that migrates to one pole. This type of division is similar to nondisjunction, in that both chromatids go to one pole. One of the resultant sperm has a B-type chromosome while the other does not. As a result, preferential fertilization can occur. Cytological results from a cross with the hemizygous telocentric showed that 0-iso disjunction was the main source of Yg bz kernels (Carlson, MNL 70: 28, 1996). A comparison of Yg bz and yg Bz kernels from this cross could give a rate of preferential fertilization for the cross. However, "nondisjunction" occurred at a low frequency, with only 29 bz kernels out of 597 (4.9%). Also, many of the bz kernels were yg in seedling phenotype, indicating 0-0 disjunction and B-9 loss at the second pollen mitosis, rather than misdivision. The yg bz class must be discarded in a test of preferential fertilization. Consequently, other crosses were examined to see if more useful genetic data could be obtained.
Perhaps coincidentally, a different method for producing unpaired type 1 telos gave a higher frequency of Yg bz and yg Bz kernel types. In this procedure, the telocentric is combined with T8-9(4453) and segregation is used to produce the constitution: 9-B 9-8 telo B-9. The B-9 is unpaired in this construct and undergoes frequent misdivision. A cross of yg yg bz bz x 9-B (Wx) 9-8 (wx) telo B-9 was made, and Wx progeny selected. Selection of Wx removes 9-8 from the data and utilizes only the 9-B B-9 meiotic product, making this cross comparable to the previous one. Classification gave a relatively high rate of bz kernels. Eight ears were classified. Among 941 Wx kernels, 77 were bz (8.1%). These kernels were grown in the field and testcrossed to analyze their constitution. The Bz kernels were classified on a sand bench for Yg vs. yg. Among 799 Bz kernels, 111 were yg. If we assume that all the bz kernels gave Yg seedlings (see later), the data show a lack of preferential fertilization because the Yg bz frequency is less than that for yg Bz. The rate of preferential fertilization calculated from these data is actually below 50%, suggesting a reversal of preferential fertilization. A calculation of preferential fertilization can be made by a) assuming that all bz kernels are Yg and b) correcting the yg Bz data for the rate of germination which is 94.7%. Preferential fertilization is then 77 divided by 77 + 117 = 40%.
The bz kernels from the cross were grown in the field, but unfortunately
they were not classified at the seedling stage for Yg vs. yg.
Therefore, the possibility of 0-0 disjunction (yg bz phenotype)
and B-9 loss could not be tested. However, similar crosses with plants
of the same genetic parentage showed no B-9 loss among 25 germinated from
bz
kernels. In any case, the existence of a yg bz class would only
reduce the rate of preferential fertilization further, because this class
would be subtracted from total bz. The conclusion is that deletion
of the centric heterochromatin, or a closely adjacent region, on type 1
telocentrics, blocks preferential fertilization. A proximal region on the
B chromosome is needed, therefore, for functioning of the preferential
fertilization system.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
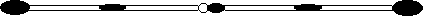{kind=link}
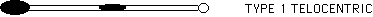{kind=link}
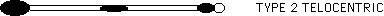{kind=link}