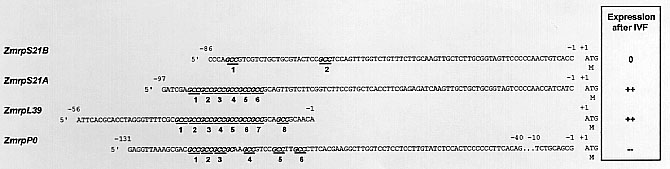
From cDNA libraries of unfertilised maize egg cells (Dresselhaus, T et al., Plant J 5:605-610, 1994) and in vitro zygotes, 18 h after gamete fusion (Dresselhaus, T et al., Plant Mol Biol 31:23-34, 1996), we have isolated more than 50 genes, whose expression is up- or downregulated after in vitro fertilisation. Among the isolated cDNAs were four encoding novel ribosomal proteins (r-proteins) from maize: ZmRPS21A, ZmRPS21B, ZmRPL39 and ZmRPP0, respectively (Accession No. X98656, Y09636, X95458 and Y07959). Relatively high transcript amounts of ZmrpS21A, ZmrpL39 and ZmrpP0 were detected in unfertilised egg cells. After in vitro fertilisation (IVF) transcript amounts of ZmrpS21A and ZmrpL39 even increased, while transcript amounts of ZmrpP0 strongly decreased (Figure 1). Transcript for ZmRPS21B was not detected in egg cells and zygotes, whereas this is the predominant transcript of the rpS21 gene family in other tissues of maize (data not shown). The homologous ribosomal genes (r-genes) from rice are regulated in a coordinated fashion during the somatic cell cycle and are strongly induced during G1 phase of the cell cycle (data not shown).
The two subunits of eukaryotic ribosomes contain more than 80 different r-proteins. Assembly of ribosomes in active nucleoli was shown to be regulated by coordinated gene expression in yeast (Mager WH and Planta RJ, Biochem Biophys Acta 949:1-15, 1988). In animals, this is different and translational controls are predominant (e.g. Aloni R et al., Mol Cell Biol 12:2203-2212, 1992). Coordinated translational control of r-protein mRNA suggests that common regulatory elements are involved. The mRNA encoding r-protein L32 from murine spp. redistributes from untranslated subribosomal particles into polysomes after mitogenic activation of quiescent cells (Kaspar RL et al., J Biol Chem 267:508-514, 1992). The regulatory element for translation was identified in the 5� UTR as a conserved polypyrimidine element. 5� UTRs of ribosomal P-proteins were shown to influence both gene expression and translation efficiency (Bermejo B et al., J Biol Chem 269:3968-3975, 1994). We have identified repetitive GCC triplets in the 5� UTRs of transcripts for maize r-proteins that are stored at relatively high amounts in unfertilised egg cells (ZmrpS21A, ZmrpL39, ZmrpP0) and which are absent in the transcripts for ZmRPS21B, of which no transcript was detected in egg cells and zygotes (Figure 1). As shown by Southern blot analysis, these repetitive triplets are very abundant in the genome of maize (Figure 2d). GCC triplets have been identified also in the human genome on chromosomes X, 11 and 16. They are polymorphic at the FRAXF site on chromosome X, where 6-29 triplets have been found (Parrish JE et al., Nat Genet 8:229-235, 1994). Extensive regions of GCC triplets were also found at the FRAXA site on chromosome X and it was shown that large triplet boxes exclude nucleosomes (Wang Y-H et al., J Mol Biol 263:511-516, 1996). In plants, GCC triplets have been described in the promoter of ethylene-induced genes. Expression is mediated by a A(GCC)2 cis-sequence element (Sato F et al., Plant Cell Physiol 37:249-255, 1996). In tobacco, gene expression was induced after virus infection mediated by an inverted GCC motif (TAAGA(GCC)2; Livne B et al., Plant Sci 130:159-169, 1997). Whether the (GCC)n-motif decribed in this paper is needed for nucleosome exclusion, gene activation or repression, transcript processing and stability, or selective mRNA translation after fertilisation remains to be determined.
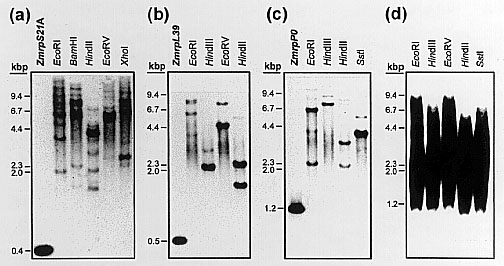
Southern blot analyses further indicated that the investigated r-genes occur as small gene families in the maize genome. We have digested genomic DNA of the maize inbred line A188 with at least four different enzymes that do not cut within the cDNA sequences. Nevertheless, due to introns within all corresponding genomic sequences (data not shown), the precise number of genes and pseudogenes cannot be predicted. In Figure 2a, seven (EcoRV) to ten (EcoRI, BamHI) bands of different signal intensities are visible after hybridization with a ZmrpS21A probe. Hybridisation with the ORF of ZmrpL39 resulted in two (HindIII) to four (EcoRI) bands (Figure 2b). A similar result was obtained with ZmrpP0, where two (SstI) to five (EcoRI, HindII) bands are visible (Figure 2c). From these data we suggest that there are probably at least two r-genes of the rpL39 and rpP0 families in the maize genome. R-genes of the rpS21 family are more abundant and at least six genes can be expected. When the full length cDNAs of ZmrpS21A or ZmrpL39 were used as probes, a smear ranging from 1.2 to 9.4 kbp was obtained (Figure 2d). The same result was achieved, when only the (GCC)8-motif of ZmrpL39 was used as hybridisation probe. Shorter exposure times of this filter revealed numerous bands (data not shown). Whether these signals are originating from repetitive (GCC)n-motifs in the maize genome or from unspecific hybridisation to GC-rich sequences has to await further experimentation, e.g. sequencing of the maize genome.
Figure 1. Fertilisation-regulated r-genes of maize contain a (GCC)n-motif in their 5� UTR. The 5� UTRs of the four r-genes presented here are aligned. GCC-triplets are numbered and printed in bold italics. The start codons are indicated with +1. The expression of the r-genes 18 h after IVF is indicated as follows: 0: expressed at undetectable levels in egg cells and zygotes, ++: strongly up-regulated and --: strongly down-regulated after IVF.
Figure
2. Genomic organisation of maize r-genes ZmrpS21A/B, ZmrpL39,
ZmrpP0
and the (GCC)n-motif. Genomic DNA was extracted from the maize inbred line
A188. Each lane contains 10 pg cDNA of the clones indicated, or 10 µg
genomic DNA digested with the enzymes shown above each lane. DNAs were
separated in 0.8% agarose gels, blotted and hybridised with probes as follows:
the ORFs of ZmrpS21A (a) and ZmrpL39 (b) were used as hybridisation
probes, respectively, the full length cDNA of ZmrpP0 was used as
probe in (c) and in (d) either the full length cDNAs of ZmrpS21A
and ZmrpL39, respectively, or the PCR amplified (GCC)8-motif of
ZmrpL39
was used as probe for hybridisation.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}