Infectious long terminal repeat (LTR) retrotransposable elements such as the animal retroviruses were once thought to be nonexistent in higher plants. Such infectious LTR retroelements (capable of moving from cell to cell) have now been postulated to exist in some higher plants. This postulated existence of infectious LTR retroelements is based on retroelement-mediated transduction of a maize cellular gene (Bureau, T, White, S, and Wessler, S, Cell 77: 479-480, 1994; Jin, Y-K, and Bennetzen, JL, Plant Cell 6: 1177-1186, 1994), and on the recent findings of conserved DNA and amino acid sequences similar to envelope (env) glycoprotein gene sequences of retroviruses as found within Arabidopsis thaliana Tat1 retroelements (Wright, DA, and Voytas, DF, Genetics 149: 703-715, 1998), and within Glycine max SIRE-1 retroelements (Laten, HM, Majumdar, A, and Gaucher, EA, PNAS 95: 6897-6902, 1998).
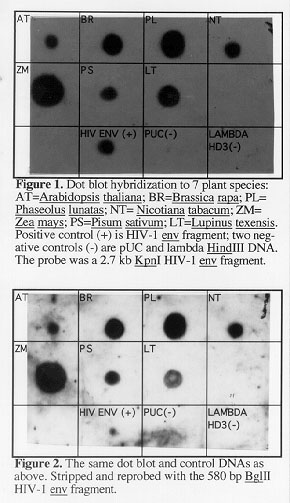
In July 1996, we obtained preliminary experimental evidence by dot blot hybridization that sequences similar to the human HIV-1 strain ADA envelope [env] gene exist in 7 higher plant species (6 dicots and 1 monocot) that were tested.
Two hybridizations were performed sequentially, both on the same Zeta probe GT nylon membrane (BioRad). Approximately 1 microgram of denatured genomic DNA from the following higher plant species was applied to the membrane: Zea mays, Brassica rapa, Arabidopsis thaliana, Nicotiana tabacum, Pisum sativum, Lupinus texensis, and Phaseolus lunatas. Then approximately 50 nanograms of denatured DNA from the following control DNAs was applied to the membrane: the probe HIV-1 env 2.7 kb gene fragment (one positive control), and pUC 18 plasmid DNA and lambda-HindIII DNA (two negative controls).
In the first hybridization (Figure 1), the dot blot was first hybridized at 35 C in aqueous hybridization solution (Ausubel et al., Curr. Prot. Mol. Biol., 1989, Wiley & Sons) overnight, with a 2.7kb HIV-1 env gene KpnI fragment obtained in 1996 from Dr. Lee Ratner of Washington University Medical School, St. Louis, Missouri (Westervelt, P, Gendelman, HE, and Ratner, L, PNAS 88: 3097-3101, 1991). This 2.7 kb KpnI fragment, which was radioactively labeled with 32P, contains almost all of the human HIV-1 env gene and a small portion of the nef gene (L. Ratner, personal communication). After hybridization and washing, the radioactive dot blot was exposed to X-ray film for 3 days (Figure 1).
In the second hybridization (Figure 2), to ensure that the positive dot blot hybridization found in the 7 plant species above was due to hybridization to the envelope gene alone, and not to the nef gene, we next probed the stripped dot blot from above with a 580 bp BglII fragment isolated from within the HIV-1 envelope gene (without any nef gene sequences) (Westervelt, P, Gendelman, HE, and Ratner, L, PNAS 88: 3097-3101, 1991). Once no radioactive signal was detectable on the dot blot after the previous, 2.7kb HIV-1 env probe hybridization, as confirmed by X-ray film exposure, we rehybridized it with the 580 bp BglII HIV-1 envelope probe at room temperature (25 C) for 3 days in aqueous hybridization solution. Following hybridization and washing, 1-day and 2-day X-ray film exposures were made. The 2-day film exposure is shown in Figure 2.
Post-hybridization washes were the same for both hybridizations, as follows: 2X SSC, 0.1% SDS, 5 min., 25 C, twice; 0.2X SSC, 0.1% SDS, 5 min., 25 C, twice; 0.2X SSC, 0.1% SDS, 15 min., 42 C, twice; 0.1X SSC, 0.1% SDS, 15 min., 60 C, twice; followed by a 2X SSC rinse.
The results in terms of presence of signals resulting from hybridization were the same for both hybridizations that were performed. As shown in Figures 1 and 2, hybridization was detected in all 7 plant species, as well as in the positive control DNA. No hybridization was detected in either of the two negative control DNAs.
Our finding in 1996 as reported here, along with other findings published in 1998 (Wright, DA, and Voytas, DF, Genetics 149: 703-715, 1998; Laten, HM, Majumdar, A, and Gaucher, EA, PNAS 95: 6897-6902, 1998), encourages us to continue our research in the following directions:
1. Identification of a wider variety of plants in which these envelope-like sequences may be present.
2. Determination of whether these sequences in plants have evolved via a vertical or horizontal transmission, and/or perhaps via horizontal transfer from animal pollinators or herbivores.
3. Determination of whether these sequences are, indeed, parts of intact and fully functional retroviruses within higher plant species.
4. If such functional, infectious retroviruses do exist in plants, attempting to understand what role(s) they may play in gene transfer and in the evolution of higher plants in general.
Acknowledgements: We wish to thank Thuy Nguyen, Jessica Golby and Anne
Hall for valuable laboratory assistance.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}