To test the hypothesis that hybridization between a wild Zea and Tripsacum (P.C. Mangelsdorf and R.G. Reeves, Texas Exp. Sta. Bull. 574, 1939; M.W. Eubanks, Econ. Bot 49:172-182 1995; M.W. Eubanks, Theor. Appl. Genet. 94:707-712, 1997) was the pivotal event in the origin of maize, DNA fingerprinting of Eastern gamagrass (Tripsacum dactyloides), perennial teosinte, (Zea diplopererennis), three species of annual teosinte (Zea mays ssp. parviglumis, Zea mays ssp. mexicana and Zea luxurians), three ancient indigenous races of maize (Nal Tel, Chapalote, and Pollo), and a modern maize inbred line (W64A) was conducted (see Table 1). The basic operating assumption of this experiment is that if maize arose via intergeneric hybridization between Tripsacum and a wild Zea, alleles shared by Tripsacum and maize that are not present in any of the teosintes should be present. To the contrary, if the phylogeny of maize is monophyletic with descent traced directly from one of the annual teosintes (G.W. Beadle, J. Hered. 30:245-247, 1939; W.C. Galinat, Ann. Rev. Genet. 5:447-478, 1971; H.H. Iltis, Science 222:886-893, 1983; J. F. Doebley, Econ. Bot. 44 (suppl):6-27, 1990), any alleles shared by maize and Tripsacum should also be found in the ancestral teosinte.
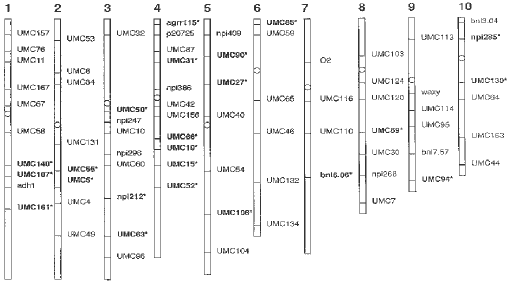
Approximately 20 grams of leaf tissue were harvested from five or more individuals of each taxa and shipped frozen on dry ice to Linkage Genetics, Inc., Salt Lake City, Utah, for DNA analysis. Methods for isolation, restriction enzyme digestion, electrophoresis, Southern transfer, labelling, and hybridization with restriction fragment length polymorphism probes are described by T. Helentjaris et al. (Pl. Mol. Biol. 5:761-769, 1985; Theor. Appl. Genet. 72:761-769, 1986). The restriction enzymes employed to digest the total bulked genomic DNA of each taxa were EcoRI, HindIII and BamHI. Proximal locations of the seventy-four molecular markers that have been mapped to the ten linkage groups of maize are indicated in Figure 1. Plant material was grown from seed in a greenhouse or obtained from clonal materials.
Analysis of the autoradiographs revealed twenty-one loci (indicated
by asterisk in Fig.
1) where Tripsacum shares alleles with one or more of the maize
lines that are not present in any of the teosintes tested. These preliminary
results lend support to the hypothesis that the evolutionary history of
maize is reticulate with descent via hybridization between Zea and
Tripsacum. An alternative explanation for these results could be
lineage sorting of alleles from a common ancestor. This seems unlikely,
however, because the frequency of >28% alleles shared between Tripsacum
and maize that are not found in teosinte is higher than would be expected
to explain these results by lineage sorting . Expanded sampling of more
taxa with more probes is needed to rigorously test the hybrid origin of
maize hypothesis. This research was supported by National Science Foundation
grant no. 9660146.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}