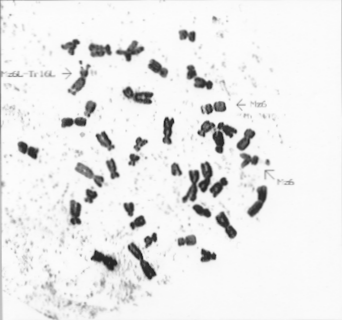
Earlier in a maize-Tripsacum hybrid line a chromosome with two satellites on both ends was discovered (Fig. 1). In maize the nucleolus-organizing chromosome is chromosome 6 and in Tripsacum chromosome 16 (Kindiger et al., Genome, 39:1133-1141, 1996). Proceeding from the morphology of the two-satellite chromosome and the lack of Tripsacum chromosome 16 in the newly-arisen line, we came to the conclusion that it was formed by a transfer of the long arm of the nucleolus-organizing gamagrass chromosome to the long arm of maize chromosome 6. The appearance of a chromosome with two satellites may be a result of meiotic crossing-over of these chromosome kinds due to available homology (Galinat, Evolution, 27:644-655, 1973; Blakey, Ph.D. dissertation, No.9412465, University of Missouri, Columbia, 1993).
In previously published results associations of maize chromosomes with Tripsacum chromosomes and mutual translocations were shown in meiosis (Maguire, Evolution, 15:394-400, 1961; de Wet and Harlan, Genetics, 78:493-502, 1974). From our data crossover exchanges between nucleolus-organizing chromosomes are also frequent enough and we found their independent emergence in three lines. Perhaps the repetition of the translocations observed by us is analogous to those described in hybrids of closely related sunflower species (Riesebery et al. Science, 272:741-745, 1996).
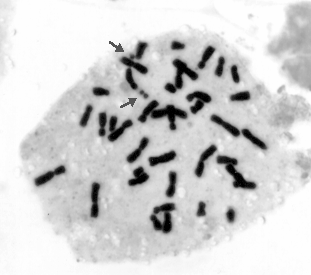
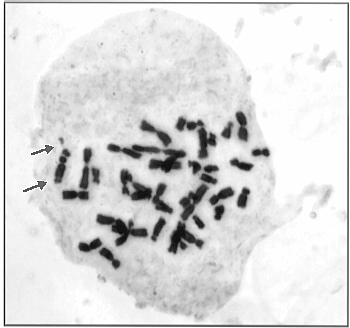
In 1997 in one plant from a line carrying a reconstructed chromosome 6MzL-16TrL we found a third satellite (Fig. 2, 3). As seen from the figures the extra (third) satellite immediately adjoins the "maize" satellite in the reconstructed chromosome. Such a phenomenon, though with two satellites only, was observed in progeny of maize-Tripsacum hybrids obtained by J. R. Harlan. The additional satellite next to that usual for chromosome 6 was found by B. Kindiger and S. A. Modena (MNL, 58:67, 1984).
We do not have experimental results explaining the mechanism of formation of the additional satellite. One can only assume a specific duplication of the nucleolus-organizing region and the satellite, or unequal crossing-over as a result of complexity of the reconstructed chromosome. Another possible explanation may be reactivation of the nucleolus-organizing region that remained quiet if adopting the hypothesis of maize tetraploid nature and the formation of chromosome 6 from two satellite chromosomes (Leitch and Bennett, Trends Plant Sci., 2(12):470-476, 1997). Other explanations are possible, however, for their adoption special experiments are needed.
For the present it is obvious that the "duplication" of the nucleolus-organizing and satellite regions has some repetition in the progeny of maize-Tripsacum hybrids and that the material from Russia carries a larger amount of satellites. The latter is in complete accordance with the high activity of their space research from the moment of the launching of the first satellite in 1957.
Acknowledgement: We are deeply thankful to Chet Dewald and B. Kindiger
for their help in preparation of the report in question.
Return to the MNL 73 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}