In the last few years, cytological evidence was advanced to demonstrate the polyploid nature of the species belonging to the genus Zea (Molina and Naranjo Theor. Appl. Genet., 73:542-550, 1987; Naranjo et al., Hereditas, 120:241-244, 1994; Poggio et al., Theor. Appl. Genet., 79:461-464, 1990; Poggio and Naranjo, Chromosome Research, 3 (Suppl.1): 80-81, 1995). These authors postulated five as the original basic number for maize and its related wild species (2n= 20) and that cryptic tetraploids originated by allopolyploidy from different diploid species (2n= 10) which are probably extinct nowadays. Some of the most suggestive evidence which allowed postulating this basic number stemmed from the analysis of the meiotic behavior of interspecific hybrids with 2n=30 chromosomes: Zea perennis x Z. diploperennis, Z. perennis x Z. mays spp. mays and Z. perennis x Z. luxurians (Molina and Naranjo, Theor. Appl. Genet. 73:542-550, 1987; Naranjo, et al., Acad. Nac. Cs. Ex. FÌs.Nat., Buenos Aires, MonografÌa 5:43-53, 1990; Poggio and Naranjo, Chromosome Research 3, Supl.1:80-81, 1995; Poggio et al., Actas XXVII Congreso Argentino de GenÈtica:185, 1996). Recent molecular approaches further confirmed the alloploid nature of maize (Moore et al., Current Biology 5: 737-739, 1995).
The genomic formula, proposed through cytogenetic studies, for taxa with 2n=20 is AxAx BxBx; for Z. perennis and hybrids with 2n=30 the formulae are ApAp A'pA'p Bp1 Bp1 Bp2 Bp2 and Ap A'p Ax Bp1 Bp2 Bx, respectively. In these hybrids the more frequent meiotic configuration was 5 III + 5 II + 5 I (Fig. 1A). Two hypotheses could be postulated to explain the original source of chromosomes in these configurations: 1) Bivalents are formed by allosyndetic pairing of chromosomes of Z. perennis and the ìBî genome of any species with 2n= 20, and the univalents would correspond to Z. perennis, i.e ApA'pAx would constitute the 5 III, Bp1Bx the 5 II and Bp2 the 5 univalents; 2) bivalents are formed by autosyndetic pairing of Bp1 and B'p2 genomes from Z. perennis and univalents would correspond to the B genome of any species with 2n= 20, i.e. ApA'pAx would constitute the 5 III, Bp1 Bp2 the 5 II, and Bx the 5 univalents.
Genomic in situ hybridization (GISH) is a molecular cytogenetic technique which now allows chromosomes from different parents or ancestors to be distinguished by means of differential hybridization of entire genomic probes (Bennett 1995, In: Kew Chromosome Conference IV, P.E. Brandham and M.D. Bennett, eds. Royal Botanic Garden, Kew, pp. 167-183 ). Using this technique in hybrids it is possible to determine the genome origin of paired and unpaired chromosomes in metaphase I configurations.
Zea luxurians has positive DAPI bands that did not hybridize with Zea perennis probe (Poggio et al., Actas XXVII Congreso Argentino de GenÈtica:185, 1996). Therefore, these chromosome regions are useful as genome markers in Z. perennis x Z. luxurians hybrids.
We present herewith the results obtained through GISH on meiotic cells of these hybrids, carried out in order to determine the origin of chromosomes forming the trivalents and univalents in the 2n= 30 Z. perennis x Z. luxurians.
Chromosome preparations: a) For meiotic configuration analysis: young panicles of the hybrids were fixed in 3:1 (absolute alcohol:glacial acetic acid). The squash of anthers was made in a drop of 2% acetic haematoxylin. b) For in situ hybridization: young panicles of the hybrid Z. perennis x Z. luxurians, growing at the IFSC, were fixed in 3:1 (absolute alcohol: acetic acid). Fixed anthers were washed in 0.01 M citric acid-sodium citrate, pH 4.6 buffer to remove fixative, transferred to an enzyme solution containing 2 ml of 2% cellulase (Onozuka R10) and 20% liquid pectinase. The softened material was again washed in the above buffer solution. Finally, anthers were squashed onto slides in a drop of 45% acetic acid. Preparations showing well spread metaphase I cells were selected by phase contrast light microscopy. After removal of the coverslip by freezing, the slides were allowed to air dry.
DNA probes: Probes consisted of total genomic DNA, which was isolated from adult leaves of the two species Zea perennis and Zea luxurians using standard methods. Probes were labelled by random priming with digoxigenin 11 -dUTP (Boehringer, Mannheim, Germany).
Fluorescent ìin situî hybridization: The technique was developed in the ìDepartamento de BiologÌa Celular y GenÈtica, Universidad de Alcal·, Alcal· de Henares, Madrid, EspaÒa) and was carried out according to Cuadrado and Jouve (Genome 38(4):795-802, 1995) with minor modifications. Slide preparations were incubated in 100ug/ml DNAse-free RNAse in 2 X SSC for 1 hour at 37 C in a humid chamber and washed three times in 2 X SSC at room temperature for 5 min. The slides were post-fixed in fresh 4% (w/v) paraformaldehyde in water for 10 min, washed in 2 X SSC for 15 min, dehydrated in a graded ethanol series and air dried.
The hybridization mixture consisted of 50% (v/v) deionized formamide 10% (w/v) dextran sulphate, 0.1% (w/v) SDS, 0.3 mg/ml of salmon sperm DNA in 2 X SSC. Then 100 ng of digoxigenin 11-dUTP probe was added to 30 ul of hybridization mixture for each slide and denatured for 15 min at 75 C. The hybridization mixture was loaded onto the slide preparation and covered with a plastic coverslip. The slides were placed on a thermocycler machine at 75 C for 7 min (denaturation), 10 min at 45 C and 10 min at 38 C. The slides were then incubated at 37 C overnight for hybridization.
Following hybridization, coverslips were carefully floated off by placing the preparations in 2 x SSC at 42 C for 3 min and then given a stringent wash in 20% formamide in 0.1 X SSC at 42 C for 10 min. The slides were washed in 0.1 X SSC at 42 C for 5 min; 2 X SSC for 5 min at 42 C; 4 X SSC 0.2% Tween 20 at 42 C for 5 min and finally for one hour in 4 X SSC/tween 0.2% at room temperature.
To detect probes, slides were treated with sheep antidigoxigenin FITC (Fluorescein isothiocyanate) . Slides were treated with 5% (w/v) BSA (bovine serum albumine) in detection buffer (4 X SSC, 0.2% (v/v) Tween-20). Then slides were incubated in a solution of the corresponding antibody in detection buffer containing 5 % BSA for one hour at 37 C and washed three times in 4 X SSC/ Tween 20 for 10 min at room temperature.
Slides were counterstained with 4¨6-diamidino-2-phenylindole (DAPI) (2 ug/ml in McIlvaine's citrate buffer, pH=7), for 10 min at room temperature and then they were mounted in antifade solution. Slides were examined with a Zeiss Axiophot epifluorescence. Photographs were taken using Fuji color super G400 color print film.
In situ hybridization was carried out on hybrid meiotic cells. Chromosomes were blocked with unlabelled Zea perennis genomic DNA and probed with digoxigenin labelled genomic DNA from Zea luxurians. This procedure allowed chromosomes from Zea luxurians to be identified by mean of their telomeric specific sequences and to be distinguished from those derived from Zea perennis, which remained unlabelled because of the blocking procedure.
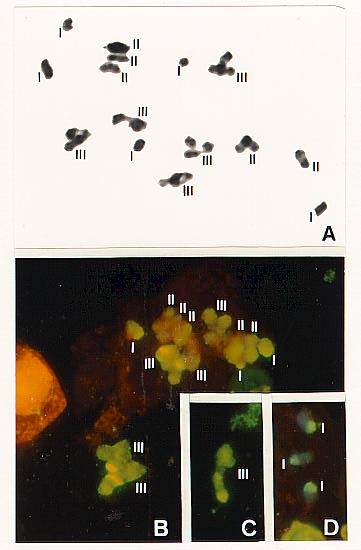
In metaphase l cells of the hybrid Z. luxurians x Z. perennis the following configurations and hybridization signals were observed: a) Univalents had a strong fluorescent signal in their telomeric regions; therefore, these chromosomes correspond to Z. luxurians, because they display the distal portion not blocked by Z. perennis DNA that bound Z. luxurians labelled DNA probe (Figs. 1B, D). b) Trivalents were composed by one chromosome labelled in its telomeric region - i.e. a Z. luxurians chromosome- and two smaller and unlabelled ones ? i.e. Z. perennis chromosomes (Figs. 1B, C). c) Bivalents did not show any label so they correspond also to Z. perennis (Fig. 1B). (ed. note: see color reproductions of Figs. 1B, C, and D in MaizeDB)
These results confirm the second hypothesis previously advanced, that in these 2n= 30 hybrids the trivalents are formed by autosyndetic pairing of Ap and A'p genomes from Zea perennis and by allosyndetic pairing with the A genomes from the tetraploid parent Z. luxurians.
Acknowledgements: We wish to express our sincere thanks to Dr. Nicolas Jouve and Angelines Cuadrado for the guidance, valuable suggestions and economical support given while L.P. and C.A.N. visited the ìDepartamento de BiologÌa Celular y GenÈtica, Alcal· de Henares, Madrid, Spainî. Financial support from the Consejo Nacional de Investigaciones Cientificas y TÈcnicas and Universidades de Buenos Aires and La Plata is also acknowledged.
Figure
1
Please Note: Notes submitted to the Maize Genetics
Cooperation Newsletter may be cited only with consent of the authors
Return to the MNL 72 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}