In native populations from northern Argentina we have found numerical polymorphism for accessory or B-chromosomes (Bs). It is reported that Bs are maintained in the populations due to various mechanisms of driving (Carlson and Roseman, Genetics 131: 211-223, 1992), consisting of: 1) Suppression of meiotic loss when they are in single doses, 2) Nondisjunction at the second pollen grain mitosis, 3) Preferential fertilization by the sperm nucleus carrying Bs produced after the nondisjunction process, 4) Competitive ability of B-carrying pollen grains.
One of the features of the Bs mode of inheritance is the variation in their transmission rate (TR) in such a way that in some progenies the Bs tend to be lost and in others they tend to increase in number compared to Mendelian expectation. The B TR was estimated by the ratio between the mean number of Bs transmitted to the progeny and the number of Bs of the parental plant that carried them. In previous papers we found that the frequency of B chromosomes in native races of maize varies considerably in different populations (Rosato et al. 1998, Am. J. Bot. 85(2), in press). Moreover, the existence of genotypes controlling high and low male transmission rate (male B-TR) of B chromosomes in Pisingallo race (Rosato et al., Am. J. Bot. 83(9):1107-1112, 1996) was demonstrated.
In the present work crosses were made to determine whether the genes controlling male B-TR are located on the normal chromosome set (A) or on the B-chromosomes. The experiment was carried out in two lines (high and low male B-TR lines) from the race Pisingallo (VAV 6313) which is a native population from the northwest of Argentina collected by the authors. We made four types of 0B female x 2B male crosses between and within both H (High) and L (Low) B-TR lines. Twenty two successful crosses were obtained as follows: (1) five H x H; (2) six H x L; (3) six L x H; and (4) five L x L (Figure 1). The number of Bs was determined in primary root tips of at least twenty individuals of each of the twenty two ears. The male B-TR values were compared with a two way ANOVA test. The frequencies of progeny with 2B, related to the frequencies of individuals with 0B, were compared with the Mendelian expectation (0.5) using a X2 test.
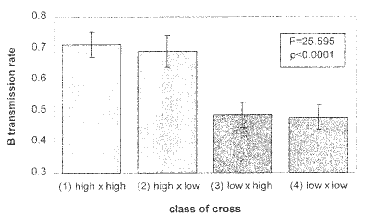
The mean B-TR of the progeny resulting from H x H crosses was = 0.71±0.04 (Table 1). The frequency of individuals with 2B was significantly higher than the Mendelian expectation (X2 =23.04, p<0.00001). The mean B-TR of the progeny resulting from L x L crosses was 0.48±0.04. The frequency of individuals with 2B did not differ significantly from the Mendelian expectation (X2=0.286, p=0.593).
The mean B-TR of the progeny resulting from H x L crosses was 0.70±0.05; this value does not differ significantly from the B-TR from H x H (p=0.99882). The frequency of individuals with 2B was also significantly higher than the Mendelian expectation (X2 = 21.893, p<0.00001). The mean male B-TR of the progeny resulting from L x H crosses was 0.48±0.04; this value does not differ significantly from the B-TR from L x L (p=0.99906). The frequency of individuals with 2B did not differ significantly from the Mendelian expectation (X2 =0.39, p<0.532).
In all cases most of the individuals were 0B or 2B, the frequency of plants with 1B or 4B being very low (Table 1).
There were significant differences in the B-TR between crosses 1 and 2 vs. 3 and 4 (Table 1), when H and L B-TR female progenitors were considered (F=26.453, p<0.0001), whereas no significant difference was found between crosses 1 and 3 vs. 2 and 4 (Table 1) when H and L B-TR male mean B-TR progenitors were considered (F=0.040, p=0.8436) (Figure 1).
Our results indicate for the Pisingallo maize population (VAV 6313) that the B-TR genotype of the receptor 0B plant determines the rate by which the egg is fertilized by the sperm nucleus carrying 2Bs. Therefore, the detection of differences between sperm with and without Bs is controlled by the female progenitor genotype. Thus, some genotypes increase the frequency of fertilization by B-carrying sperm nucleus, whereas other genotypes do not. The latter result in random fertilization by the 0B or 2B sperm nucleus, leading to a Mendelian B-TR. All these results indicate that the presence of B-chromosomes determines the sperm phenotype which would be accepted by the egg cell. In addition, since in our experimental design the female progenitor has no Bs, we conclude that these controlling genes are located on the regular A chromosome set.
Table 1. Types of crosses with their male B transmission rate (male
B-TR). L: low male B-TR line, H: high male B-TR (data from Chiavarino et
al., Am. J. Bot. 85, 1998, accepted).
| type of cross | no. of | no. of indiv. with different no. of Bs | mean B-TR | type of | |||
| (female x male) | crosses | 0B | 2B | 4B | total | ± SE | B-TR |
| 1) 0B (H) x 2B (H) | 5 | 42 | 99 | 1 | 142 | 0.71±0.04 | high B-TR |
| 2) 0B (H) x 2B (L) | 6 | 50 | 109 | 2 | 161 | 0.70±0.05 | high B-TR |
| 3) 0B (L) x 2B (H) | 6 | 86 | 78 | 1 | 165 | 0.48±0.04 | Mendelian |
| 4) 0B (L) x 2B (L) | 5 | 66 | 60 | 0 | 126 | 0.48±0.04 | Mendelian |
Figure
1. Types of crosses with their male B transmission rate. low: low male
B-TR line, high: high male B-TR.
Return to the MNL 72 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}