Partial isolation and characterization of the maize alternative oxidase
gene
--Polidoros, AN, Mylona, PV, Scandalios, JG
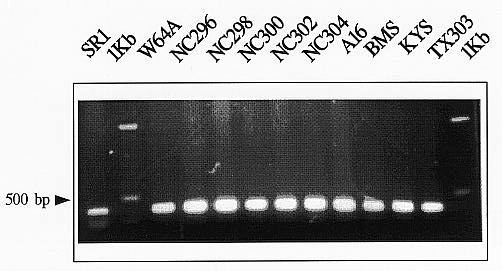
Plant mitochondria contain an alternative pathway for the transfer of electrons from reduced ubiquinone to oxygen that bypasses cytochrome c oxidase and is associated with the presence of an alternative oxidase found in the inner membrane of mitochondria from all higher plants. Recent evidence suggests that alternative oxidase has a potentially important role in protection against oxidative stress. In order to characterize the maize alternative oxidase, we amplified part of the gene with PCR, and DNA template from different maize inbred lines (Figure 1), using degenerate primers designed from two highly conserved regions of alternative oxidase from other plant species. The degenerate primer specific to the 5Ì conserved region was 5Ì-GCIATGATGYTIGARACIGTRCG-3Ì and corresponds to the conserved amino acid sequence AMMLETVA. The degenerate primer specific to the 3Ì end of the gene was 5Ì-RTCICGRTGRTGWGCCTCRT-3Ì and its complementary strand encodes the conserved amino acid sequence DEAHHRD. PCR products of the expected size (450bp in the tobacco SR1 control) were observed on an agarose gel with all the templates tested (Figure 1). The PCR bands were subcloned and these of the maize lines NC300 (Goodman M., Proc. Annu. Corn Sorghum Ind. Res. Conf. 47:47-64, 1992), W64A, A16 and the tobacco line SR1 were sequenced. The sequence derived from the tobacco line was identical with the respective region of the alternative oxidase sequence from SR1 (X79768) in the GenBank. The sequence derived from the maize line W64A had no similarity with other alternative oxidase sequences and may represent a pseudogene or misprimed amplification. The sequences from NC300 and A16 were very similar and that of NC300 was selected for further analysis. The NC300 PCR product had 81.8% identity with the respective nucleotide sequence of S. guttatum (M60330), and 87.2% amino acid identity, as was revealed by sequence alignments (Figure 2). Using the cloned fragment of the maize alternative oxidase gene derived from the line NC300 as probe, we performed RFLP analysis with four inbred maize lines. Results indicated that the gene is polymorphic among the genotypes examined (data not shown).
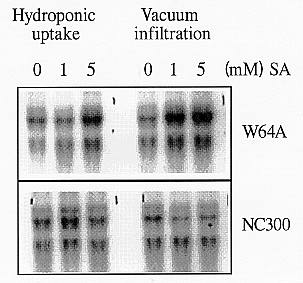
Alternative oxidase in other organisms is inducible by salicylic acid (SA). Thus, we examined if the maize alternative oxidase gene is also inducible by SA. We treated 10 dpi W64A and NC300 maize seedlings with 1 and 5 mM SA, either hydroponically or with vacuum infiltration. After 24 h treatment we examined the effects on alternative oxidase transcripts. The results showed that multiple transcripts were observed and each genotype responded differently to SA (Figure 3). In all cases three bands were observed. The upper band was weak and did not seem to alter with treatments. The middle and lower bands were clearly responding, with the responses of the middle one more prominent. The middle band corresponds to the expected alternative oxidase transcript size if the maize transcript is similar with that of other plants. The W64A alternative oxidase transcripts were highly increased in the vacuum infiltrated seedlings with both 1 and 5 mM SA, when in the hydroponic treatment they increased only in the higher SA concentration. However the NC300 alternative oxidase transcripts increased only in the 1 mM SA hydroponic treatment. In the vacuum infiltration treatments the middle band seemed to decrease. Using the PCR-derived maize alternative oxidase probe we plan to isolate the cDNA and genomic DNA of the gene and characterize its responses to oxidative stress.
Figure 1. Electrophoretic analysis of PCR products. PCR was performed using a set of degenerate primers, which amplify part of the maize alternative oxidase gene, and template DNA from the indicated maize lines and the tobacco line SR1. PCR conditions were 3 min denaturation at 950C followed by 35 cycles of 30sec denaturation at 950C, 1min annealing at 580C, and 1min extension at 750C. The last extension step was for 5 min to ensure completion of the reaction. A single band of fairly similar size and intensity can be observed.
Figure 2. Alignment of the alternative oxidase nucleotide sequence (A) and amino acid sequence (B) from S. guttatum and the maize NC300 PCR product. The maize nucleotide sequence has been submitted to the GenBank and accession number assignment is AF040566. Different nucleotides and amino acids are highlighted in the maize sequences.
Figure 3. Alternative oxidase inducibility with SA. Northern analysis of RNA (20µg) isolated from 10 dpi W64A and NC300 maize seedlings treated with 1 and 5 mM SA either hydroponically or with vacuum infiltration. The NC300 derived alternative oxidase probe was used in this experiment. Note the 3 bands clearly detected with the probe. The upper band is fairly constant in all the lanes. The middle band is the most intense and corresponds to the expected size of the alternative oxidase transcript. Different induction is observed between the two lines.
Return to the MNL 72 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}